Il ruolo del DNA nell'ereditarietà - Gli esperimenti di Fredrick Griffith su Streptococcus pneumoniae hanno dimostrato la presenza di un ...
←
→
Trascrizione del contenuto della pagina
Se il tuo browser non visualizza correttamente la pagina, ti preghiamo di leggere il contenuto della pagina quaggiù
Il ruolo del DNA nell’ereditarietà Gli esperimenti di Fredrick Griffith su Streptococcus pneumoniae hanno dimostrato la presenza di un «principio trasformante» ereditabile.
Il ruolo del DNA nell’ereditarietà Alfred Hershey e Martha Chase (nel 1952) scelsero un virus, il batteriofago T2, per determinare quale dei componenti virali (DNA o proteine) sarebbe penetrato nel batterio Escherichia coli. Tali scienziati dimostrarono che il materiale con ruolo ereditario è il DNA; esso penetra nelle cellule batteriche e trasmette ai batteri infettati le informazioni genetiche necessarie a produrre nuovi virus.
Il ruolo del DNA nell’ereditarietà
Gli esperimenti di Hershey e Chase con i batteriofagi T2 ed E. coli.
I
II
vedi sopra
3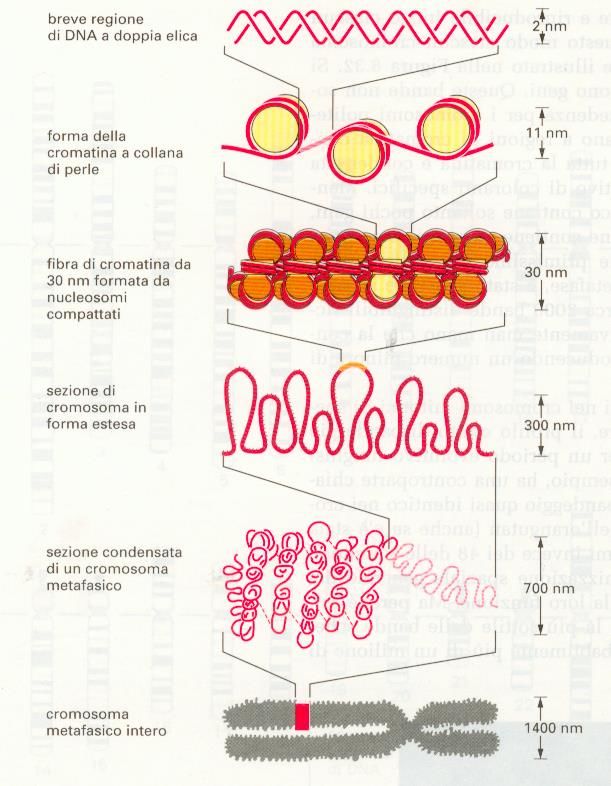
LA STABILITA’ DELLA MOLECOLA DI DNA A
DOPPIA ELICA E’ ASSAI ELEVATA
DOVUTA A TRE ORDINI DI FATTORI
PRESENZA DEGLI SCHELETRI COVALENTI
DELLE DUE CATENE COMPLEMENTARI
INTERAZIONI IDROFOBICHE TRA COPPIE
SOVRAPPOSTE DI BASI
LEGAMI A IDROGENO ALL’INTERNO DI OGNI
COPPIA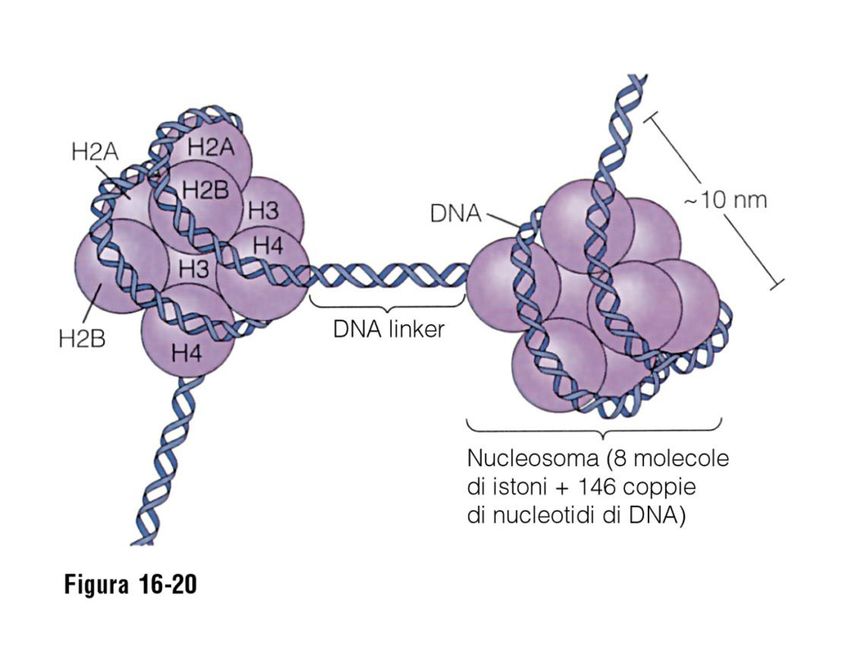
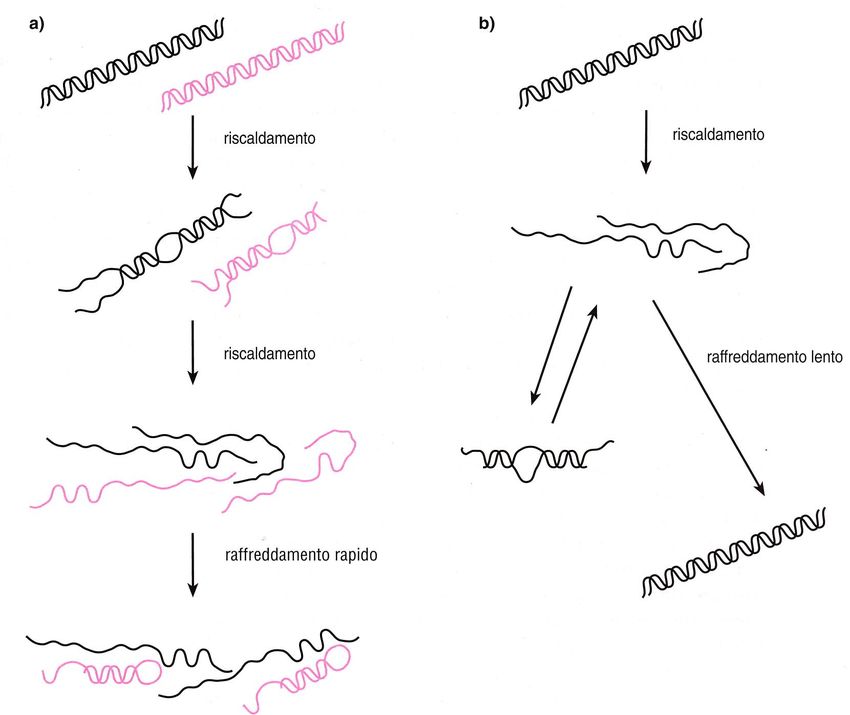
Denaturazione e rinaturazione del DNA

Il DNA e l’RNA, gli acidi nucleici,
sono polimeri di nucleotidi
Il DNA è un polinucleotide, cioè
un polimero di nucleotidi, ognuno
formato da tre parti:
• uno zucchero C5 detto
desossiribosio;
• un gruppo fosfato;
• una base azotata.
6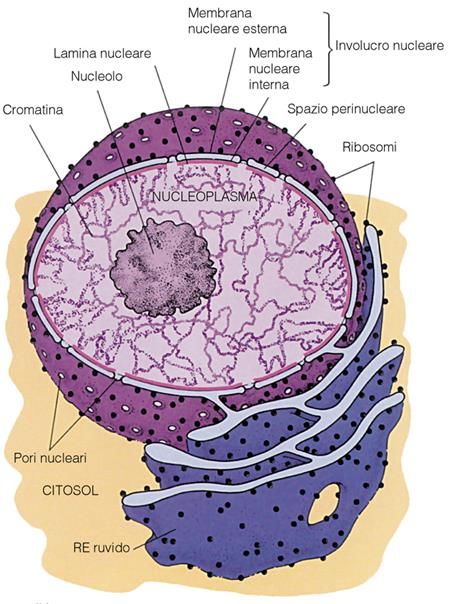
Il DNA e l’RNA, gli acidi nucleici, sono
polimeri di nucleotidi
Esistono quattro diversi tipi di basi azotate. L’adenina (A) e la
guanina (G) sono caratterizzate da un doppio anello e sono chiamate
purine. La timina (T) e la citosina (C) sono caratterizzate da un
anello singolo e sono chiamate pirimidine.
7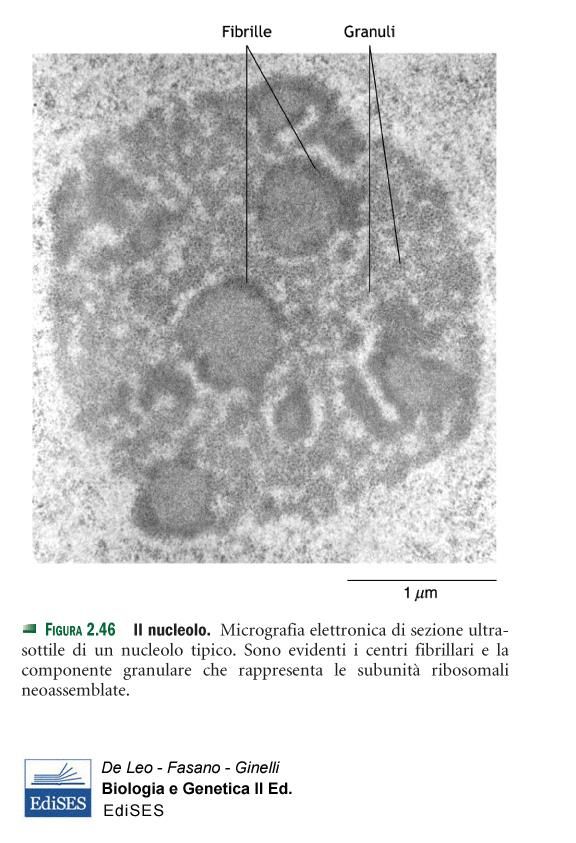
Il DNA e l’RNA, gli acidi nucleici, sono
polimeri di nucleotidi
L’RNA (acido ribonucleico) differisce dal DNA per il tipo di
zucchero C5 che contiene, il ribosio, e perché al posto della base
azotata timina contiene un’altra pirimidina, l’uracile (U).
8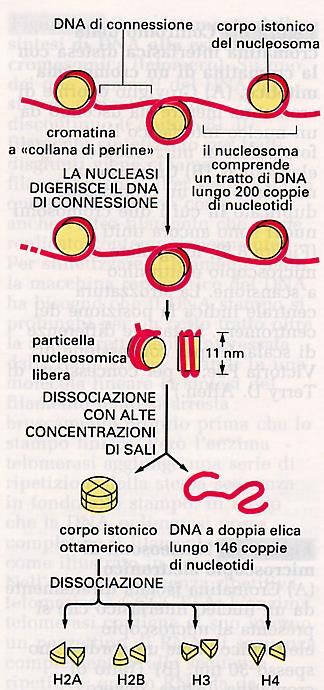
Il DNA ha i requisiti adatti per funzionare come materiale genetico • Il DNA è variabile tra le diverse specie. • Il DNA è in grado di custodire le informazioni che fanno una specie diversa dall’altra. • Il DNA è costante all’interno di una stessa specie. • Il DNA è in grado di duplicarsi con grande precisione durante la divisione cellulare. • Il DNA è soggetto a rari cambiamenti, chiamati mutazioni, che forniscono la variabilità genetica che permette agli organismi di evolversi nel tempo.
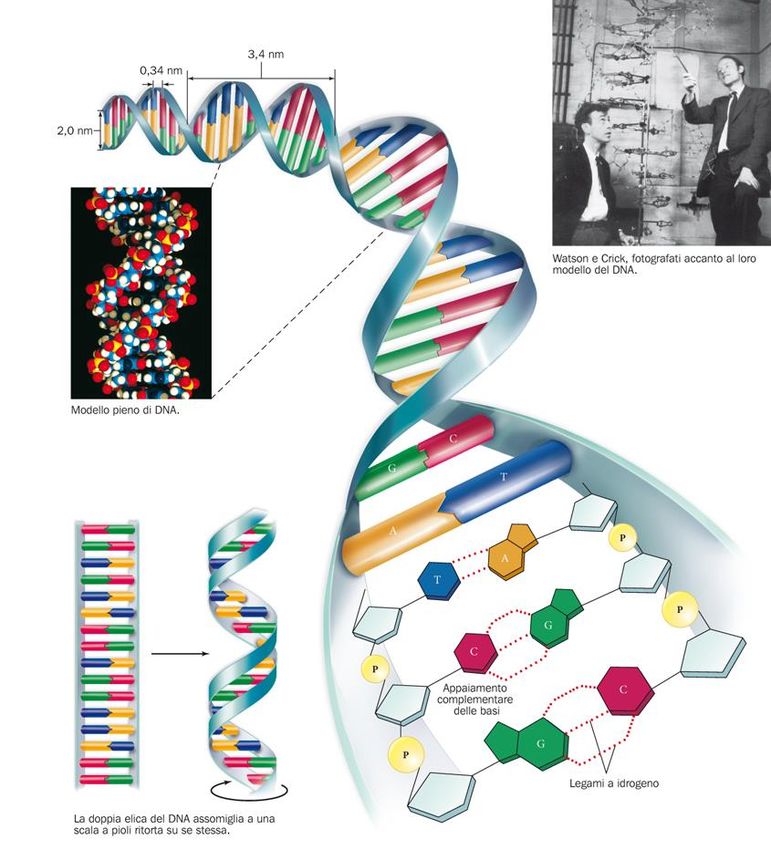
La struttura del DNA La molecola del DNA ha la forma di una doppia elica di nucleotidi in cui la base A è sempre in coppia con la base T, e la base C con la base G. 10
La molecola del DNA ha la forma di una
doppia elica
James Watson e Francis Crick costruirono il primo modello
tridimensionale del DNA basandosi sui risultati dei lavori di
Rosalind Franklin e Maurice Wilkins, che avevano studiato la
struttura del DNA usando la cristallografia a raggi X.
11La molecola del DNA ha la forma di una doppia elica L’appaiamento complementare delle basi azotate suggerisce che il DNA è una molecola a doppio filamento, simile a una scala a pioli in cui i montanti sono costituiti dallo scheletro zucchero- fosfato, e i pioli dalle basi accoppiate unite da legami idrogeno.
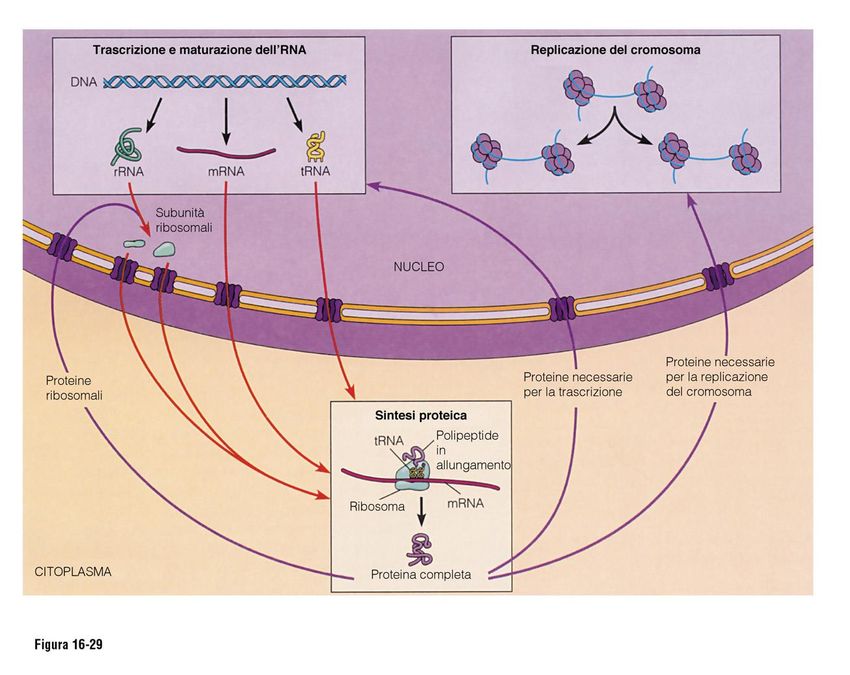
Il nucleo interfasico
Aspetto del materiale genetico
Di una cellula in interfaseNUCLEO
E’ L’ORGANULO PIÙ
VOLUMINOSO DELLA CELLULA
EUCARIOTICA
SOLITAMENTE APPARE DI
FORMA ROTONDEGGIANTE E
FREQUENTEMENTE
LOCALIZZATO NELLA REGIONE
CENTRALE DEL CITOPLASMA
LA MAGGIOR PARTE DEGLI EUCARIOTI PRESENTA
UN SOLO NUCLEOCOSTITUISCE LA SEDE
DELL’INFORMAZIONE
GENETICA: IN ESSO
RISIEDONO LE MOLECOLE
DI DNA CHE VENGONO
DUPLICATE E TRASCRITTE
CROMATINA
CROMOSOMI
TERRITORIO CHE DELIMITA
LE ATTIVITA’ DEL GENOMA
(INSIEME DI GENI DI UNA
CELLULA) E PROTEGGE DNA
ed RNA DALL’AMBIENTE
CITOPLASMATICOIL NUCLEO È UNA
PORZIONE DEL
CITOPLASMA DELIMITATO
DA UN SISTEMA DOPPIO DI
MEMBRANE CHE PRENDE IL
NOME DI
INVOLUCRO NUCLEARE o
CARIOTECA E CHE
RACCHIUDE IL
NUCLEOPLASMAMEMBRANA NUCLEARE
INTERNA e MEMBRANA
NUCLEARE ESTERNA
SEPARATE DA UNO
SPAZIO PERINUCLEARE
OGNI MEMBRANA HA LA
CARATTERISTICA STRUTTURA
BILAYER DELLA MEMBRANA
CELLULARE.
LA MEMBRANA NUCLEARE
ESTERNA È IN CONTINUITA’
CON IL RETICOLO
ENDOPLASMATICO RUGOSO.LA MEMBRANA NUCLEARE
INTERNA È POGGIATA SU
UNA SOTTILE E DENSA RETE
DI PROTEINE ALTAMENTE
INSOLUBILI, ADESA AL
VERSANTE
CITOPLASMATICO DELLA
MEMBRANA INTERNA DELLA
CARIOTECA, CHIAMATA
LAMINA NUCLEARE.
LAMÌNE NUCLEARI A, B, C
FUNZIONE
• SOSTEGNO
• SITO DI ATTACCO PER I TELOMERI DEI CROMOSOMI DURANTE
LA MEIOSI
• DIVISIONE NUCLEARE (MITOSI, MEIOSI): LA FOSFORILAZIONE
DELLE LAMÌNE DETERMINA LA FRAMMENTAZIONE
DELL’INVOLUCRO NUCLEAREPORI NUCLEARI: METTONO IN COMUNICAZIONE IL
COMPARTO NUCLEARE CON QUELLO CITOPLASMATICO
COMPLESSO DEL PORO
NUCLEARE (NCP)ORGANIZZAZIONE OTTAGONALE: OTTO PROTEINE POSTE IN
MANIERA SIMMETRICA ATTORNO AL PORO CHE FORMANO
DUE ANELLI CONCENTRICI APPOGGIATI ALLE DUE
MEMBRANE
OTTO RAGGI CHE
DIPARTONO DAGLI ANELLI E
RAGGIUNGONO IL
TRASPORTATORE POSTO AL
CENTRO DEL PORO
PROTEINE CHE SI
ESTENDONO DAL BORDO
VERSO LO SPAZIO
PERINUCLEARE
(ANCORAGGIO DEL
COMPLESSO)OTTO FIBRE CHE SI ESTENDONO DAGLI ANELLI VERSO IL
CITOSOL E VERSO IL NUCLEOPLASMA (SONO UNITE ALLA
LORO ESTREMITA’ DA UN ANELLO FIBROSO)
NUCLEOPORINE
CLASSE PIU’ ABBONDANTE
DI PROTEINE PRESENTI NEL
NUCLEO
TRASPORTO PASSIVO (PM = <
15000 Da)
TRASPORTO ATTIVO (PM =
15000 Da) E SELETTIVOBIOGENESI DEI RIBOSOMI
MATRICE NUCLEARE
FIBROGRANULARE
INSOLUBILE
(NUCLEOSCHELETRO)
FUNZIONE
• FORMA DEL NUCLEO
• IMPALCATURA PER
L’ORGANIZZAZIONE DELLE
FIBRE DI CROMATINA
• SISTEMA DI ANCORAGGIO
PER COMPLESSI
MOLECOLARI COINVOLTI
NELLA DUPLICAZIONE E
NELLA TRASCRIZIONEL’analisi chimica dimostra nel nucleo la presenza di DNA , RNA e
proteine, a questo complesso viene dato il nome di
CROMATINA: complesso di DNA, proteine
(ISTONI E NON ISTONICHE) ed RNA
DNA GENOMICO: PIU’ MOLECOLE DI DNA
DNA DEL NUCLEO DI UNA CELLULA UMANA: 1.7 METRI
DIAMETRO DEL NUCLEO: 0.5 MICRONIl materiale genetico subisce una
Transizione da fibrillare a bastoncellare.
Da una organizzazione idonea a
Permettere al materiale genetico di essere
Trascritto e reduplicato, a quella idonea
Di essere parimenti distribuito tra
Le cellule figlie
Poro nucleare
MetafaseAspetto della Cromatina a collana di perle dopo trattamento con soluzioni saline
NUCLEOSOMA
1) PARTICELLA CENTRALE: COMPLESSO DI OTTO PROTEINE
ISTONICHE – DUE MOLECOLE DEGLI ISTONI H2A, H2B, H3 E
H4
2) DNA A DOPPIO FILAMENTO, LUNGO 146 COPPIE DI
NUCLEOTIDI.
L’ OTTAMERO
DI ISTONI
FORMA UN
NUCLEO
PROTEICO
INTORNO AL
QUALE E’
AVVOLTO IL
DNA A DOPPIO
FILAMENTOOrganizzazione del NUCLEOSOMA Il trattamento della cromatina con DNAsi, Permette di isolare una struttura contenete DNA e proteine, la lunghezza minima che si Riesce ad ottenere con tale trattamento è di 146 paia di basi che ha l’aspetto di una perla. Ogni “perla” è costituita da un nocciolo Proteico intorno al quale si avvolge il DNA. Il nocciolo proteico più il DNA che si avvolge intorno costituiscono il NUCLEOSOMA
MODELLO A “COLLANA DI PERLE”
CIASCUNA PARTICELLA
CENTRALE DEL NUCLEOSOMA E’
SEPARATA DALLA SUCCESSIVA DA
UNA REGIONE DI DNA LINKER. LA
FORMAZIONE DEI NUCLEOSOMI
CONVERTE UNA MOLECOLA DI DNA
IN UN FILO DI CROMATINA LUNGO
CIRCA UN TERZO DELLA
LUNGHEZZA INIZIALE E CIO’
COSTITUISCE IL PRIMO LIVELLO
DI COMPATTAMENTO DEL DNAGRUPPO DI PROTEINE ETEROGENEO,
SPECIALIZZATO IN:
• DUPLICAZIONE DNA (DNA POLIMERASI,
ELICASI, GIRASI, PRIMASI…)
• TRASCRIZIONE DI GENI (RNA POLIMERASI)
• MATURAZIONE DEL PRE-mRNA
• REGOLAZIONE DEL PROCESSO DI
TRASCRIZIONE DI SPECIFICI GENI
Mobilità elettroforetica delle varie
classi di Istoni.Alcune proprietà delle proteine istoniche -Le proteine istoniche sono ricche in Lisina ed Arginina, sono proteine basiche dotate di una carica netta positiva. -Sono proteine altamente conservate.
LA CROMATINA DI UNA CELLULA VIVENTE
RARAMENTE ADOTTA LA FORMA ESTESA DI
“COLLANA DI PERLE”
I NUCLEOSOMI SONO COMPATTATI L’UNO
SULL’ALTRO GENERANDO SCHIERE REGOLARI IN
CUI IL DNA E’ ANCORA PIU’ CONDENSATO.
COSI’ LA MAGGIOR PARTE DELLA CROMATINA SI
PRESENTA SOTTO FORMA DI UNA FIBRA CON UN
DIAMETRO DI CIRCA 30nm, (FIBRA A SOLENOIDE)
CONSIDEREVOLMENTE PIU’ SPESSA DELLA
CROMATINA SOTTO FORMA DI “PERLINE SU UN
FILO”.PRESENZA DELL’ISTONE H1, PIU’ GRANDE DEGLI ALTRI
ISTONI E MENO CONSERVATO
L’ISTONE H1 SI SISTEMA SUL ROCCHETTO DI
AVVOLGIMENTO, “PINZETTANDO” IL DNA IN ENTRATA E
USCITA DAL ROCCHETTO E AVVICINANDO I DIVERSI
ROCCHETTI GRAZIE A LEGAMI DI TIPO TESTA-CODA
CON ALTRI ISTONI H1COME FIBRA DI 30nm UN TIPICO CROMOSOMA UMANO
SAREBBE ANCORA LUNGO 0.1cm.
CI DEVE ESSERE UN LIVELLO ANCORA SUPERIORE DI
RIPIEGAMENTO CHE ORGANIZZA LA FIBRA DI 30 nm IN
UNA SERIE DI ANSE E AVVOLGIMENTI“SCAFFOLD” PROTEICO
NONOSTANTE L’ALTO GRADO DI COMPATTAMENTO
DELLA CROMATINA LA SUA STRUTTURA DEVE
ESSERE ALTAMENTE DINAMICA PER PERMETTERE
ALLA CELLULA DI SVOLGERE LE SUE FUNZIONI
(CRESCERE E DIVIDERSI)
A QUESTO SCOPO SONO IMPORTANTI DUE STRATEGIE
GENERALI PER CAMBIARE REVERSIBILMENTE
STRUTTURE LOCALI DI CROMATINA:
•COMPLESSI DI RIMODELLAMENTO DELLA
CROMATINA, SPINTI DA ATP
• MODIFICAZIONE COVALENTE (ACETILAZIONE,
METILAZIONE, FOSFORILAZIONE) CATALIZZATA
DEGLI ISTONI (H1)CICLO DI CONDENSAZIONE-DECONDENSAZIONE DELLA
CROMATINAA SECONDA DELLE SUE ATTIVITÀ TRASCRIZIONALI LA
CROMATINA PUÒ ESSERE FORTEMENTE ADDENSATA E
QUINDI INATTIVA (ETEROCROMATINA) O LASSAMENTE
COMPATTATA E QUINDI ATTIVA (EUCROMATINA).
•ETEROCROMATINA: COSTITUTIVA (RESTA ALLO STATO
COMPATTATO SEMPRE E IN TUTTE LE CELLULE, NON
VIENE MAI TRASCRITTA); FACOLTATIVA (RISULTA
INATTIVATA IN MODO SPECIFICO IN ALCUNE FASI DELLA
VITA DI UN ORGANISMO)
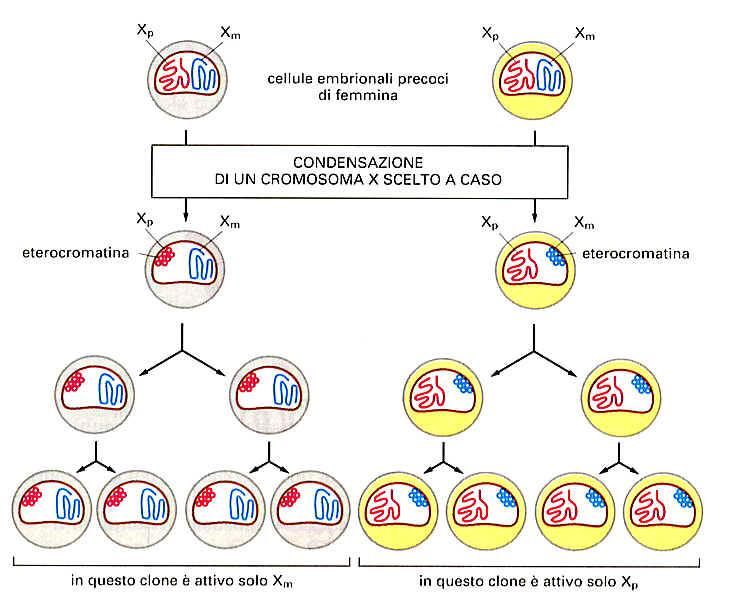
ES: SI HAIN UNO DEI DUE CROMOSOMI X INTERFASICI
DELLE FEMMINE DI MAMMIFERI: LE LORO CELLULE
POSSIEDONO DUE CROMOSOMI X, MENTRE QUELLE DEI
MASCHI HANNO UN X E UN Y.NEI MAMMIFERI UNO DEI CROMOSOMI X FEMMINILI VIENE INATTIVATO
PERMANENTEMENTE, FORSE PERCHE’ UNA DOPPIA DOSE DI CERTI PRODOTTI
GENICI POTREBBE ESSERE LETALE. ALLORA UNO O L’ALTRO DEI DUE, A CASO, SI
ADDENSA FORTEMENTE E DIVENTA ETEROCROMATICO IN UNA FASE PRECOCE
DELLO SVILUPPO EMBRIONALE.
DA QUEL
MOMENTO, IN
TUTTA LA
NUMEROSA
PROGENIE
CELLULARE CHE
SEGUE, LO STATO
DI
CONDENSAZIONE
E INATTIVITA’ DI
QUEL
CROMOSOMA X
VIENE
EREDITATO.CROMOSOMI
STRUTTURA MOLTO COMPATTA E COLORABILE, VISIBILE
AL MICROSCOPIO OTTICO DURANTE LA DIVISIONE
CELLULARE, IN PARTICOLARE DURANTE LA METAFASE,
QUANDO VIENE RAGGIUNTO IL MASSIMO LIVELLO DI
COMPATTAMENTO DELLA CROMATINA.
Ogni cromosoma è una
lunga molecola di DNA
(assieme a specifiche
proteine) la quale
contiene le informazioni
necessarie al
funzionamento della
cellula (e che verranno
trasmesse alle cellule
figlie)In ogni specie i cromosomi si presentano con un numero e una
struttura caratteristici. Ad esempio il corredo cromosomico umano è di
46 cromosomi (23 coppie di cromosomi omologhi).
Tutte le cellule di un dato organismo
hanno lo stesso numero di cromosomi
(ad eccezione dei gameti che ne
hanno esattamente la metà)CROMOSOMA MITOTICO
METACENTRICI
SUBMETACENTRICI o
ACROCENTRICI
TELOCENTRICI
BRACCI
Cromosoma non duplicato
CROMOSOMICI
BRACCIO CORTO (p)
BRACCIO LUNGO (q)
BANDEGGIO COESINE
REGIONI e BANDE
(numeri arabi dal
centromero verso la parte Cromosoma duplicato
terminale del cromosoma)
14q2.1NUCLEOLO
STRUTTURA SPECIALIZZATA COMPRENDENTE UN
GRUPPO DI GENI CHE PORTANO L’INFORMAZIONE PER
GLI RNA RIBOSOMALI (rDNA), I TRASCRITTI
CORRISPONDENTI E PROTEINE.REGIONE FIBRILLARE:
contiene l’rDNA (ORGANIZZATORE
NUCLEOLARE) IN FORMA DI
CROMATINA PARZIALMENTE
COMPATTA, RNA POLIMERASI I E
FATTORI DI TRASCRIZIONE.
I centri fibrillari sono circondati da una
COMPONENTE FIBRILLARE
DENSA = geni ribosomiali
trascrizionalmente attivi → cromatina
meno condensata
REGIONE GRANULARE:
particelle ribosomali a vari stadi di
assemblaggio
LUOGO DI ASSEMBLAGGIO DELLE
SUBUNITÀ DEI RIBOSOMIPuoi anche leggere

















































