Cellule procariotiche - Eubatteri Archeobatteri - Infermieristica
←
→
Trascrizione del contenuto della pagina
Se il tuo browser non visualizza correttamente la pagina, ti preghiamo di leggere il contenuto della pagina quaggiù
Cellule procariotiche Eubatteri Archeobatteri
I batteri
Organismi unicellulari di piccole dimensioni (1-10mm), possono associarsi
in filamenti o formare piccoli ammassi di cellule
Il genoma varia da 0,5 a 5
milioni di coppie di basi ed è
in grado di codificare tra
1000 a 4000 proteine
differenti
Hanno strutture membranose
interne molto ridotte
La popolazione batterica evolve
in fretta e aquisisce rapidamente
la capacità di utilizzare una
nuova fonte di sostentamento o
di resistenza agli effetti letali di
un nuovo antibiotico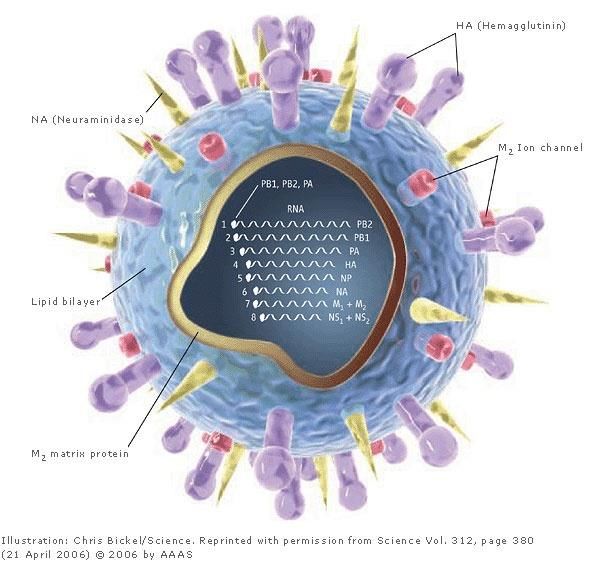
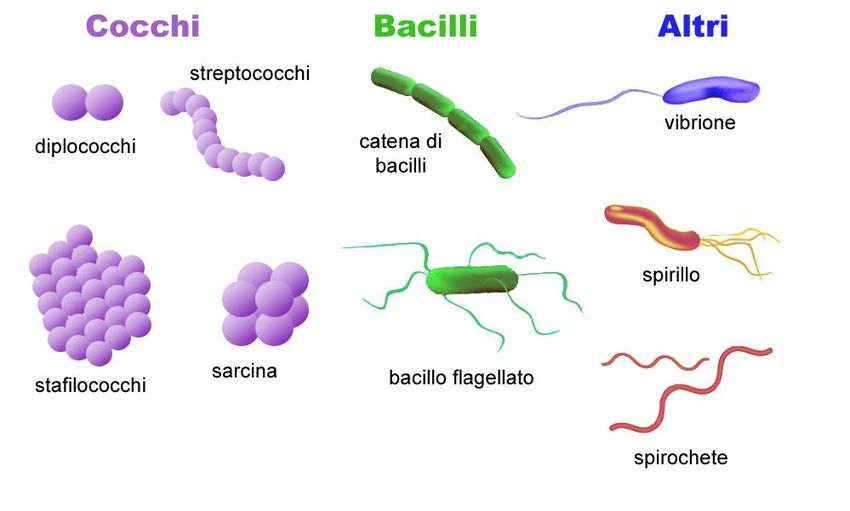
I procarioti presentano forme diverse:
I cocchi sono cellule di forma approssimativamente sferica: possono presentarsi
come singola sfera oppure riunite in aggregati
Strutture tridimensionali
o agglomerati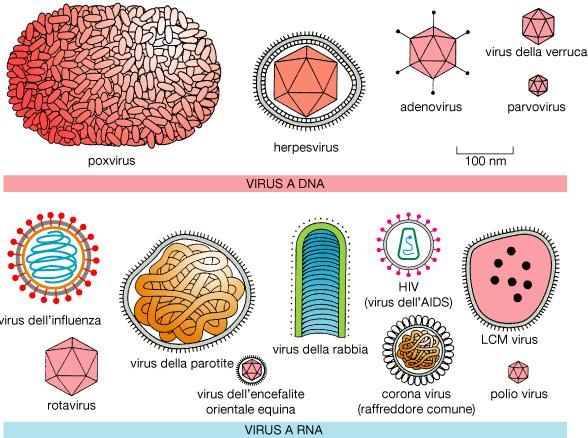
Forma dei batteri: Cocchi
i cocchi restano uniti
in coppia al
momento della
divisione
Divisione casuale
Formazione di lunghe catene I cocchi si dividono su tre Gruppi di forma quadrata
piano, formando strutture
cubiche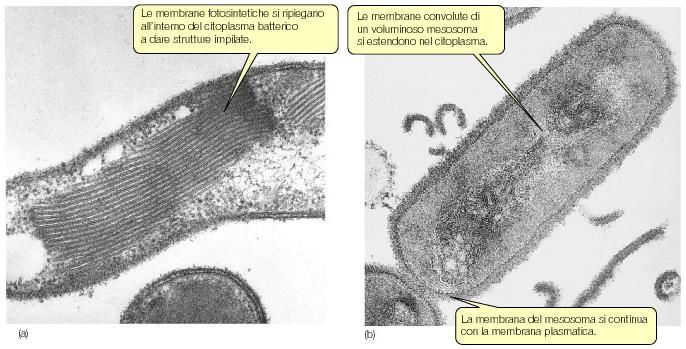
Forma dei batteri: bastoncelli o bacillo
I bacilli differiscono notevolmente rispetto al rapporto lunghezza/larghezza
Le estremità del bastoncello spesso
differiscono: si possono avere
estremità piatte, arrotondate, a forma
di sigaro o biforcute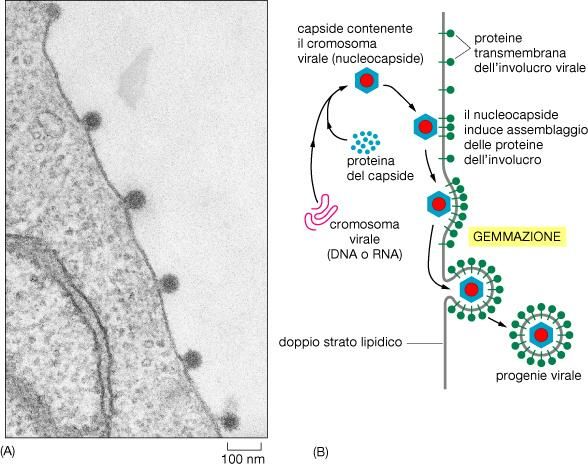
I batteri
Le principali strutture dei batteri
sono:
-Parete cellulare
-Membrana plasmatica
-Ribosomi
-Inclusioni citoplasmatiche (materiale
di riserva fatto di composti a base di
carbonio, azoto, zolfo o fosforo)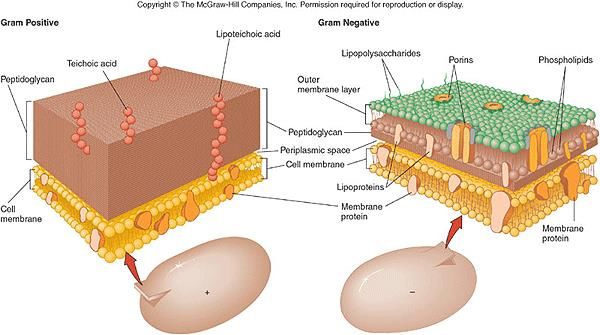
La parete cellulare
La parete cellulare è una struttura che circonda la cellula all’esterno della
membrana plasmatica
Composta principalmente peptidoglicano
-Mantenimento della forma
-Conferimento della forza meccanica
-Controllo dell’espansione
Gram+ Gram-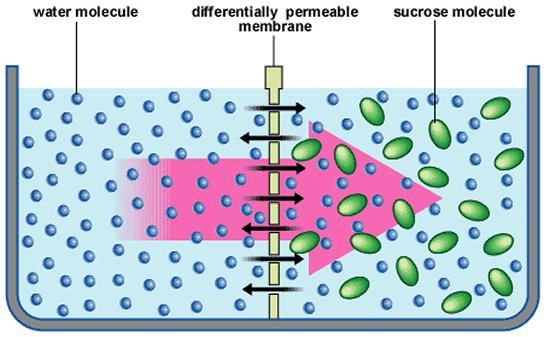
Colorazione di Gram
La colorazione di
Gram è il più importante
metodo di colorazione batterica
differenziale, definita così perché
non colora in maniera uguale
tutti i tipi di cellule.
Il preparato contenente i
batteri viene trattato con:
-cristalvioletto
- mordenzatore (soluzione di
iodio e ioduro di potassio in
acqua) una sostanza che
permetta al colorante di
legarsi in maniera stabile al
substrato
- decolorante (alcol etilico o
acetone)
- colorante che può essere la
fuxina o la sufranina
-di decoloranti.Gram-positivi: i batteri appaiono Gram-negativi: i batteri sono
colorati in violetto perché il decolorati e pertanto assumono
decolorante non è riuscito a la colorazione del secondo
rimuovere il complesso colorante utilizzato, quindi
cristalvioletto-iodio da queste cellule appaiono rossi
L’idrofobicità della parete cellulare è alla base della caratteristica Gram-positività: mentre il
cristalvioletto e lo iodio veicolato dalla soluzione di Lugol riescono a penetrare nella cellula
attraversando la parete cellulare grazie alla loro idrofilia, il complesso colorante-iodio che si
forma all’interno della cellula assume caratteristiche idrofobiche che non gli consentono di
riattraversare la parete cellulare per essere estratto dalla cellula anche in presenza di
decoloranti.I batteri
Parete cellulare: differenza tra Gram positivi e Gram negativi
Strato rigido formato da peptidoglicano sottile lamina costituita da due derivati
polisaccaridici e da un gruppo di amminoacidi assemblati a formare unità di
ripetizione, il glican-tetrapeptide
Acidi lipoteicoici
Carica -
Acidi teicoici
Carica -
Il peptido glicano
rappresente solo il
Il peptidoglicano
10% della parete,
rappresente fino al
90% del materiale di
parete
Spazio periplasmatico rete di peptidoglicano a Lo strato esterno (lipopolisaccaride) è costituito da
maglie larghe enzimi coinvolti nei processi di
assorbimento dei nutrienti, nella sintesi del
lipopolisaccaride contenete fosfolipidi, polisaccaridi e proteine
peptidoglicano e nella modificazione dei Complesso lipoproteico svolge funzioni di ancoraggio tra membrane
composti tossici esterna e peptidoglicano
Proprietà biologica: tossicità per gli animaliLa parete cellulare e la protezione osmotica Osmosi: l’acqua si muove attraverso le membrane selettivamente permeabili passando da soluzioni diluite a soluzioni più concentrate
Lisi: l’acqua entra nella cellula, la pressione osmotica aumenta e determina la lisi della cellula Plasmolisi: negli abitat ipertonici fuoriuscita dell’acqua, il citoplasma si raggrinzisce e si stacca dalla parete cellulare (meccanismo utile nella conservazione dei cibi)
La membrana plasmatica La membrana plasmatica è una barriera selettiva costituita da lipidi nei quali sono inglobate alcune proteine
Membrana plasmatica un sistema altamente
organizzato e asimmetrico:
-proteine periferiche legate alla membrana da legami deboli 20-30%
-proteine integrali associate al doppio strato lipidico 70-80%
- Trattiene il citoplasma
- Barriera selettiva permeabile
- Assunzione di nutrienti
- Eliminazione di cataboliti
- Secrezione di proteine
- Respirazione
- Fotosintesi
- Sintesi dei lipidi e dei costituenti della parete
- Segregazione cromosomica
-Alcuni procarioti dispongono di complessi
sistemi di membrane interne
Membrane fotosintetiche mesosomi
•mesosomi settali: intervengono nella divisione cellulare, legano il cromosoma batterico e
producono il setto trasversoLa matrice citoplasmatica sostanza compresa
fra la membrana plasmatica e il nucleoide
- costituita dal 70% di acqua
- altamente organizzata e ricca
di ribosomi
Proteine specifiche sono
localizzate in regioni particolari
della cellula
Protoplasto: membrana
plasmatica e tutto ciò che in
essa è contenutoIl nucleoide
Le cellule procariotiche non possiedono un nucleo
L’informazione genetica (aploide) è racchiusa in un’unica molecola,
cromosoma procariotico situato in una regione di forma irregolare,
chiamata, nucleoide
Il nucleoide è in contatto con la membrana
plasmatica o con il mesosoma
Formato da:
60% DNA
30%RNA
10%proteine (diverse da quelle
eucariotiche)
I plasmidi molecole di DNA circolare
- Crescita cellulare
- Riproduzione dell’ospite
- Vantaggi selettivi
- Resistenza a farmaci
- Nuove capacità riproduttive
L’assenza di membrana nucleare fa sì che
una molecola di mRNA venga tradotta nella
proteina corrispondente mentre procede
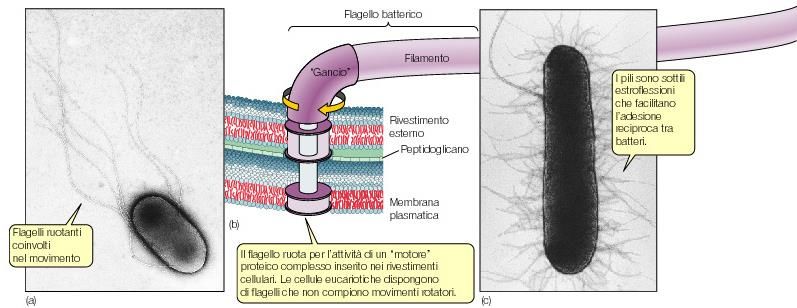
ancora la sua trasrizioneStrutture accessorie dei batteri
• Fimbrie o pili: brevi appendici filiformi, sono sottili tubuli, formati da
subunità proteiche organizzate in una struttura elicoidale
– Permettono ai microorganismi di aderire a superfici inerti o di formare pellicole
o strati sulla superficie di sostanze liquide
– Motilità contrattile
– -sono coinvolti nel processo di coniugazione batterica meccanismo di
trasferimento genetico
adesione
coniugazioneStrutture accessorie dei batteri
Flagelli appendici cellulari lunghe e sottili libere ad un’estremità costituiti da
una singola proteina la flagellina. Il flagello batterico è una struttura rigida ed
elicoidale, che si inserisce alla base della cellula e ruota a spese del gradiente di
protoni determinando il movimento cellulare
flagellinaLe appendici delle cellule procariotiche
L’endospora batterica
Molti batteri gram positivi sono in grado di generare al loro
interno una struttura quiescente e particolarmente resistente
a stress ambientali come calore radiazioni ultraviolette,
radiazioni gamma, disinfettanti chimici e disidratazione
Le specie batteriche che le producono
sono pericolosi patogeniVIRUS
1890 Iwanosky e Beijerinck
Estratto ottenuto da piante di tabacco affette da malattia
“del mosaico” – agente infettivo filtrabile
1935-1960 Struttura dei virus
Microscopia elettronica
CARATTERISTICHE PECULIARI
- Entità infettive contenenti acido nucleico (DNA o RNA), che si replicano
all’interno di cellule viventi utilizzano l’apparato biosintetico della cellula
per dirigere la sintesi di particelle virali (VIRIONI)
- Diametro dei virioni varia tra 10 e 300-400nm
- I virioni NON crescono di dimensioni e NON si dividono. Vengono
MONTATI per interazione di macromolecole
- I virioni non hanno metabolismo e sistemi enzimatici per la produzione
di energia
- I virioni necessitano di cellule infettate per riprodursiSTRUTTURA DEI VIRIONI -Acido nucleico (sempre presente) -Capside (sempre presente) -Involucro membranoso (presente in alcuni virus animali) Il virione è metabolicamente inerte e non esplica funzioni respiratorie e biosintetiche. Il processo attraverso cui un genoma virale si introduce e si replica in una cellula viene chiamato infezione La cellula in cui un virus può penetrare e riprodursi viene detta ospite Il virus riprogramma quelle componenti metaboliche e biosintatiche preesistenti nell’ospite, che sono necessarie per la sua replicazione
Proprietà strutturali generali
Nucleocapside: acido nucleico, DNA o RNA, racchiuso da un involucro
formato da proteine organizzate in maniera precisa e molto ripetitiva
intorno all’acido nucleico
tipi di capside con conformazioni altamente simmetriche:
a) capside Icosaedrico (solido regolare a 20 facce triangolari)
b) capside elicoidali (forma simile a cilindri proteici cavi, rigidi o
flessibili
c) Virioni a struttura complessa (i. senza un vero e proprio capside; ii.
con capside non icosaedrico o tubulare, iii. con capside
asimmetrico per la presenza di strutture accessorie come code,
fibre etc.)
-Involucro membranosoCapside: duplice funzione protegge l’acido nucleico e
permette l’interazione del virus con la cellula infettata
Struttura a forma sferica
La geometria icosaedrica
rappresenta la disposizione
più efficiente per le
subunità di un capside,
perchè utilizza il minor
numero di capsomeri per
costruire il rivestimentoStruttura del virus del mosaico del
tabacco
Virus ad RNA con simmetria elicoidale in cui le
subunità proteiche identiche sono disposte ad
elica.
La lunghezza dei virus elicoidali è determinata
dalla lunghezza del genoma, mentre la
larghezza dal grado di impacchettamento delle
subunità proteicheVirus a struttura complessa
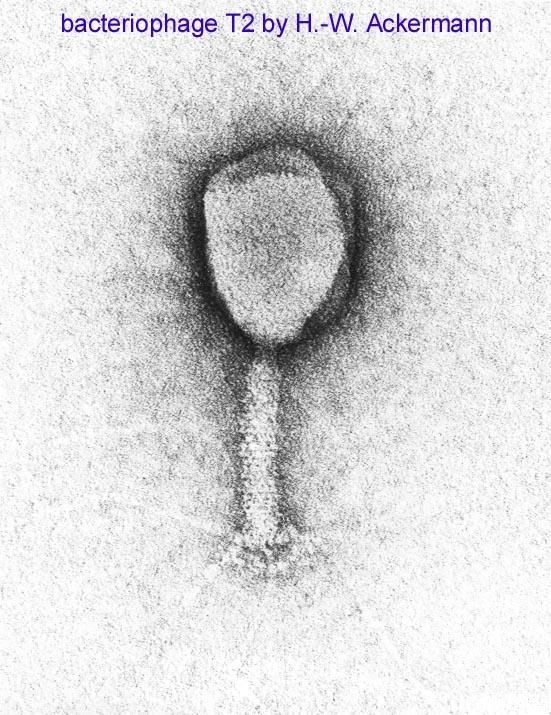
Virioni composti da diverse parti distinte ciascuna con la sua
configurazione e simmetria
Testa icosaedriche
T4
Batteriofago lambda Coda elicoidale
Con fibre caudaliAcidi nucleici
- DNA o RNA
- A doppio o singolo filamento
- Lineare o circolare che può propagarsi da un ospite ad un altro
- Intero o segmentato
-Virus influenza: 8 segmenti di RNA a singolo filamento
-Retrovirus: 2 molecole di RNA a singolo filamento (diploide)
-Dimensione variabile
- 2.000-200.000 basi (5-50% peso del nucleocapside)
-Geni sovrapposti
-cornici di lettura sovrapposte
-geni più piccoli contenuti in geni più grandiAcidi nucleici RNA a doppio filamento
STRUTTURA DEI VIRIONI
-Acido nucleico
-Capside
-Involucro membranoso o pericapside doppio strato lipidico contenente
glicoproteine
Caratteristico di alcuni virus animali. Proviene dalla membrana
plasmatica o da altre membrane della cellula ospite (RE, Nucleo,
Golgi) modificate con proteine virali (dette spicole).
Esempio: virus dell’influenza.
-HA=spicole di emoagglutinina per l’attacco del virione alla
superficie
della cellula ospite
-NA= spicole di neuroaminidasi per la liberazione dei nuovi virioni
dalla cellula ospiteCaratteri generali
Virioni: Sferici, oppure filamentosi
(pleomorfici)
Involucro: Lipidico, contiene 2
glicoproteine HAe NA con uno strato
costituito di 2 proteine di matrice M1
e M2
Genoma: RNA a polarità negativa a
unico filamento segmentato(8
segmenti in A e B, 7 in C)
Core: Nucleoproteina associata ad
ogni filamento di RNA formando i
ribonucleoproteine (RNPs)
Simmetria: Elicoidale
Replicazione: Nucleare
Virus dell’influenzaEnzimi dei virus: -DNA polierasi RNA dipendente, retrotrascrive l’informazione genetica dell’RNA genomico in un intermedio ad DNA -neuroaminidasi, rompe i legami glicosidici delle glicoproteine e dei glicolipidi del tessuto connettivo aiutando la liberazione della progenie virale. -lisozima, enzima con attività battericidi, nei batteri Gram + catalizza l'idrolisi del legame beta 1,4 tra l'acido N-acetilmuramico (NAM) e la N- acetilglucosamina (NAG) che sono la componente principale del peptidoglicano producendo un foro nella parete batterica
TROPISMO VIRALE -Generalmente un virus può infettare un numero ristretto di tipi cellulari; ciò fornisce una base per classificarli in: Virus batterici (batteriofagi), Virus animali e Virus vegetali -Alcuni virus però possono infettare sia piante sia insetti che si cibano di esse (Esempio: un virus della patata infetta anche locuste che si nutrono delle foglie della pianta di patata) -Alcuni virus animali possiedono una specificità ampia (esempio: virus della stomatite vescicolare che infetta insetti e molti tipi di cellule di mammifero) -La maggior parte dei virus, però, sono specifici per phyla -Alcuni virus sono estremamente specifici e infettano solo poche specie o addirittura solo alcuni tipi cellulari -La specificità è dovuta all’interazione fra proteine virali presenti sulla superficie del capside e specifiche strutture esposte sula superficie della cellula infettate,
Classificazione dei virus animali Data la limitatezza del materiale genetico disponibile, il tipo di geni presenti nei genomi virali è ridotto al minimo necessario per consentire: a) la replicazione del DNA o RNA virale b) La formazione del capside
Esempi di virus animali Osservazioni: I virus variano molto sia per dimensioni che per forma e non tutti i virus hanno l’involucro esterno
Caratteristiche generali della replicazione dei virus I vari aspetti del processo di replicazione di un virus possono essere suddivisi in sette fasi: -Attacco (adsorbimento) del virione ad una cellula ospite sensibile -Penetrazione (iniezione) del virione o del suo genoma all’interno della cellula -Fasi replicative precoci del genoma virale, in cui il macchinario biosintetico della cellula ospite è modificato, come preludio alla sintesi di acidi nucleici virali, vengono sintetizzati enzimi virus-specifici -Replicazione del genoma virale -Sintesi delle subunità proteiche -Assemblaggio delle subunità proteiche del capside virale -Assemblaggio delle subunità proteiche e impacchettamento del genoma nelle nuove particelle virali -Rilascio di virioni maturi dalla cellula (lisi)
Infezione dell’ospite da parte di un
batteriofago
Adsorbimento del fago T4 alla parete cellulare di E.Coli
-Attacco delle lunghe fibre
caudali alla parete cellulare
-Contatto fra la parete
cellulare e la punta della coda
-Azione di un enzima
lisozimico che permette la
formazione di un foro
-Contrazione della guaina e
iniezione del DNAAdsorbimento e penetrazione della cellula ospite Adesione dei batteriofagi alla superficie della cellula ospite in corrispondenza di specifiche strutture, recettori Adsorbimento: le fibre caudali giungono a contatto con il recettore, la piastra basale si dispone sulla superficie Pistra e guaina si riorganizzano formando un cilindro Penetrazione dell’asse tubulare centrale all’interno della parete batterica (gp5 proprietà lisozimatiche) Il DNA è estruso dalla testa, attraversa l’asse tubulare caudale ed è introdotto nella cellula ospite
Sintesi degli acidi nucleici
e delle proteine fagiche
- la sintesi di proteine, DNA e RNA dell’ospite è interrotta
- RNA polimerasi dell’ospite viene indirizza a sintetizzare
mRNA fagici
- Alcuni enzimi precoci virali virus-specifici degradano il DNA
dell’ospite per ricavarne nucleotidi
- Sintesi DNA virale e proteine fagiche del capside
- Assemblaggio del virus maturoLiberazione delle particelle fagiche
Ciclo litico di un batteriofago
Endolisina: attacca il peptidoglicano della
parete cellulare
Olina: lesione della membrana plasmatica
che arresta la respirazioneCiclo litico e ciclo lisogeno I virus temperati possono entrare in uno stato, detto di lisogenia, nel quale non viene espressa la maggior parte dei geni fagici, e il genoma virale si replica in sintonia con il cromosoma dell’ospite Viene espressa solo una proteina cI che funge da repressore di tutti gli altri geni virali
Lisi o lisogenia?
Il genoma di lambda contiene due gruppi
di geni, uno per il controllo della crescita
litica e l’altro per il controllo della
lisogenia
La competizione fra questi prodotti genici
precoci e l’influenza di fattori dell’ospite
determina quale di queste vie avrà
sucesso
Dipende dallo stato nutrizionale della cellula ospite.
I geni fagici sono mantenuti stabilmente nello stato lisògeno finchè non
avviene un cambiamento per il quale passano, con grande efficienza, in
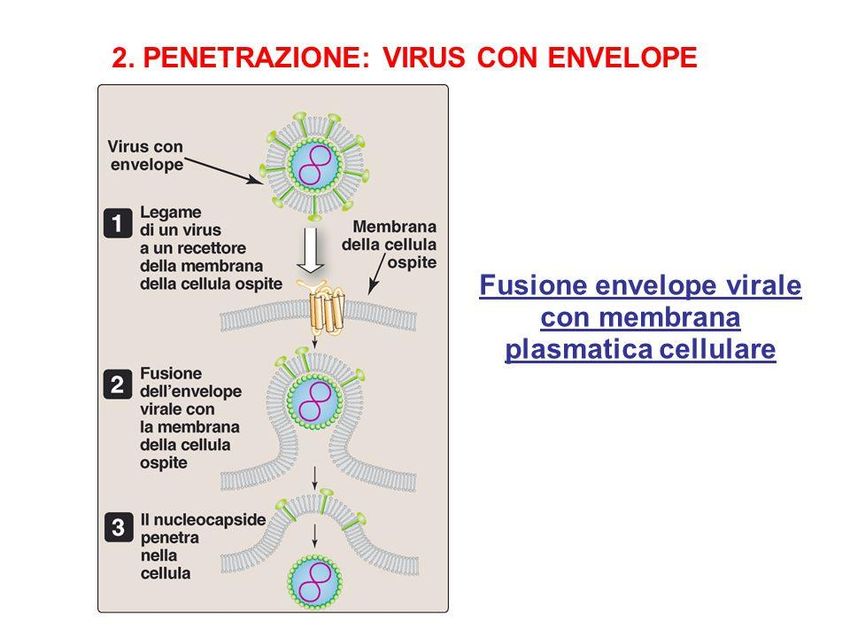
un secondo stato in cui si verifia la crescita liticaCiclo di un virus animale 1. Adsorbimento del virus alla cellula Recettori specificità 2. Ingresso nella cellula - Inglobamento del virione (fagocitosi o endocitosi) - Fusione dell’involucro virale con la membrana cellulare 3. Liberazione dell’acido nucleico -citoplasma -nucleo 4. Trascrizione del genoma virale 5. Traduzione -Poliproteine Proteine virali 6. Replicazione dell’acido nucleico 7. Assemblaggio nuovi virioni 8. Liberazione delle particelle virali - Lisi cellulare (virus senza involucro) - Gemmazione (virus con involucro) - Esocitosi (virus con involucro)
Ciclo di un virus animale a DNA
Gemmazione dei virus animali Proteine virali dell’involucro attraversano la membrana e si legano a proteine del capside Nel caso di virus con involucro il rilascio di virioni, che avviene con un processo di gemmazione, può essere lento e la cellula può non essere lisata, rimanere viva e continuare a produrre virus per un lungo periodo di tempo
Gemmazione dei virus animali
Acquisizione
dell’involucro tramite
gemmazione
Gemmazione del virus del morbilloCiclo di un retrovirus
I retrovirus sono virus a RNA che si replicano a DNA
Genoma diploide:due molecole di RNA a singolo filamento tenute
insieme da legami idrogeno per mezzo di specifiche molecole di tRNA
Processi di replicazione di un retrovirus:
-Ingresso nella cellula
-Retrotrascrizione di una delle due subunità
genomiche di RNA in un DNA a singolo
filamento, che è succccessivamente
convertito in un DNA lineare a doppio
filamento, per mezzo della trascrittasi inversa
-Integrazione della copia a DNA nel genoma
dell’ospite
-Trascrizione del DNA virale, che porta alla
formazione degli mRNA virali e dell’RNA
genomico virale
-Incapsidazione dell’RNA virale in
nucleocapsidi nel citoplasma
-Gemmazione dei virioni rivestiti attraverso
membrana citoplasmatica e rilascio dalla
cellulaCiclo di un retrovirus
CANCEROGENESI VIRALE All'inizio degli anni '60 i virologi hanno dimostrato che: – Il fenotipo tumorale poteva essere conferito a cellule normali in coltura in seguito ad infezione con un virus (Dulbecco, 1963) – Questi virus erano capaci di integrarsi nel genoma cellulare tramite la trascrittasi inversa (Temin; Baltimora, 1970)
Virus tumorali (oncogèni) I virus che possiedono la caratteristica di indurre alterazioni dei normali processi omeostatici del ritmo proliferativo cellulare, vengono definiti virus oncogeni, a loro volta distinti in virus oncogeni con genoma a DNA e virus oncogeni a RNA - Alcuni tumori sono associati all’infezione da parte di virus oncogeni - Infatti alcuni virus contengono oncogeni, un gene che potenzialmente trasforma le cellule normali in cellule cancerose • Quando l’oncogene è trasmesso da virus: ONCOGENE VIRALE
Come si definisce un virus tumorale?
• Presenza di parte del genoma virale nei tumori ed espressione di
alcuni geni virali.
• L’infezione di cellule in vitro porta alla trasformazione
Saggi di tumorigenicita’:
• Crescita in basso siero (ridotta richiesta di fattori di crescita)
• Crescita in soft agar (crescita indipendente da ancoraggio)
•Identificazione dei geni virali che trasformano le cellule in coltura
• l’infezione in un modello animale induce tumore
– non possibile nell’ uomo
– la vaccinazione previene l’insorgenza del tumoreVIRUS ONCOGENI
• virus oncogeni a RNA (retrovirus)
– retrovirus: carcinogenesi rapida
carcinogenesi lenta
• virus oncogeni a DNA (oncoDNAvirus)
– hepadnavirus
– papovavirus
– adenovirus
– herpesvirus
– poxvirusVirus tumorali a DNA
Cellule permissive: replicano il virus e subiscono l’effetto citopatogeno, in
conseguenza del quale esse muoiono nel momento in cui rilasciano i virioni >
esse non diventano cellule tumorali
Cellule non permissive: non replicano il virus infettante e subiscono la
trasformazione neoplastica in quanto si verificano alcuni eventi molecolari atti a
determinare il fenotipo tumorale (mutagenesi inserzionale)
In una piccola percentuale di queste cellule il cromosoma virale si integra nel
genoma dove viene replicato insieme al genoma della cellula ospite o forma un
plasmide che si replica in maniera controllata.
Cambiamenti genetici nella cellula ospite
Trasformazione neoplasticaVIRUS ONCOGENI A DNA La trasformazione si verifica in assenza di moltiplicazione virale in cellule non- permissive e si realizza sempre attraverso l'integrazione del virus nel genoma della cellula ospite • L'integrazione avviene a caso e la probabilità che dia trasformazione è molto bassa VIRUS ONCOGENI A DNA • Il virus dell’epatite B (HBV) ed epative C (HCV) frequentemente associato alla comparsa del carcinoma epatico • Il virus di Epstein-Barr (EBV) associato alla comparsa del Linfoma di Burkitt, oltre che agente eziologico della mononucleosi infettiva • I papilloma virus umani (HPV) di cui diversi ceppi sono associati a carcinomi della cervice uterina
Virus tumorali a RNA
Come portano ad una alterazione genetica permanente?
Trascrittasi inversa e integrasi
Retrovirus!VIRUS ONCOGENI AD RNA
• I retrovirus sono così chiamati perché nel genoma è presente un gene (pol)
che codifica per l’enzima virale trascrittasi inversa (reverse transcriptase, RT)
che, una volta avvenuta la penetrazione del virus all’interno della cellula,
trasforma l’RNA del genoma virale in DNA, che viene incorporato nel genoma
cellulare
• Trasformanti acuti > rapida comparsa dei tumori, trasformazione
neoplastica di cellule normali in coltura e sono portatori di oncogeni
• Trasformanti lenti > sono privi di oncogeni, lungo periodo di
incubazione e non sono capaci di indurre trasformazione neoplastica
di cellule normali in coltura
• I retrovirus oncogeni responsabili di carcinogenesi rapida provocano delle
vere e proprie forme acute di tumore (soprattutto sarcomi) che si
manifestano nel giro di pochi giorni
• Il virus del sarcoma di pollo scoperto da Rous nel 1911 è il più rapido dei
virus oncogeni: i fibroblasti in coltura acquistano il fenotipo trasformato
dopo 24 ore dall'infezione
• i tumori da essi provocati sono di origine policlonaleVIRUS ONCOGENI AD RNA • I retrovirus oncogeni responsabili di carcinogenesi lenta provocano tumore solo dopo settimane o mesi (anni nel caso di retrovirus umani) dall'infezione • I tumori da essi provocati sono di origine monoclonale • HTLV-1 e HTLV-2 = human T cell leukemia virus sono associati alla comparsa nell’uomo di una particolare forma di leucemia a cellule T e di linfomi • I retrovirus HIV-1 e HIV-2 (agenti eziologici della sindrome da immunodeficienza acquisita) non sono considerati virus oncogeni; essi però causando uno stato di grave immunodeficienza, favoriscono la comparsa di tumori (kaposi’s sarcoma) che sono definiti tumori opportunistici
Puoi anche leggere

















































