EFFETTI DEI CAMBIAMENTI CLIMATICI SULLA DEMOGRAFIA DELLA FAUNA SELVATICA: LA DINAMICA DEL CERVO NELLE ALPI COZIE - POLITesi
←
→
Trascrizione del contenuto della pagina
Se il tuo browser non visualizza correttamente la pagina, ti preghiamo di leggere il contenuto della pagina quaggiù
SCUOLA DI INGEGNERIA CIVILE AMBIENTALE E TERRITORIALE CORSO DI LAUREA IN INGEGNERIA PER L'AMBIENTE E IL TERRITORIO EFFETTI DEI CAMBIAMENTI CLIMATICI SULLA DEMOGRAFIA DELLA FAUNA SELVATICA: LA DINAMICA DEL CERVO NELLE ALPI COZIE Relatore: Prof. Paco Melià Tesi di laurea magistrale di: Andrea Scialabba Matricola: 858294 Anno Accademico 2016–2017
II
Sommario I territori alpini di alta quota sono luoghi ricchi di biodiversità e sono molto sensibili ai cambiamenti climatici: mostrano, infatti, un tasso di aumento delle temperature superiore alla media globale. La risposta più evidente delle specie alpine ai cambiamenti climatici è uno spostamento delle popolazioni verso quote più elevate. Le specie che occupano quote maggiori sono quindi più vulnerabili perché l'habitat disponibile si restringe all'aumentare della quota. Lo studio della demografia della fauna selvatica alpina e della sua dipendenza dalle principali variabili ambientali è quindi cruciale per valutare la vulnerabilità di queste specie nel prossimo futuro. In questo lavoro di tesi sono stati sviluppati modelli di dinamica di popolazione per tre popolazioni di cervo (Cervus elaphus) nelle Alpi Cozie italiane, con l'obiettivo di indagare il ruolo del cambiamento climatico sulla dinamica demografica di questa specie. A tale scopo sono stati utilizzati i dati di censimenti condotti a partire dagli anni ‘80 in tre Parchi (Parco del Gran Bosco di Salbertrand, Parco Orsiera-Rocciavrè e Parco della Val Troncea) e forniti dall'Ente di Gestione dei Parchi delle Alpi Cozie. È stato testato un ampio insieme di modelli dinamici in cui i parametri demografici sono stati espressi in funzione della densità e dei principali fattori meteorologici (temperatura, precipitazioni, altezza del manto nevoso). L'identificazione dei modelli migliori (in termini di capacità predittive e di parsimonia) è stata effettuata utilizzando due diversi criteri di model selection (l’Aikake’s Information Criterion e il Bayesian Information Criterion). I modelli selezionati incorporano il ruolo dell’altezza della neve e della precipitazione cumulata nell’influenzare le dinamiche della specie. Questi fattori ambientali, insieme alla densità di popolazione, interagiscono nel determinare la demografia di tutte e tre le popolazioni, nonostante la presenza di alcune importanti differenze tra una popolazione e l'altra. I modelli calibrati tramite la ricostruzione degli andamenti storici delle tre popolazioni sono stati poi utilizzati per predire la dinamica demografica futura mediante simulazione stocastica (Monte Carlo). I modelli sono stati forzati tramite serie temporali sintetiche delle variabili ambientali corrispondenti a diversi possibili scenari di cambiamento climatico (in particolare, gli scenari RCP 4.5 e RCP 8.5 prodotti dall’IPCC). I risultati delle simulazioni in avanti non evidenziano rischi rilevanti per due delle tre popolazioni (Gran Bosco di Salbertrand e Orsiera-Rocciavrè). Per la terza (Val Troncea), invece, le previsioni ottenute con entrambi gli scenari indicano un livello di rischio elevato. È opportuno notare, tuttavia, che il modello relativo alla popolazione della Val Troncea è stato ricavato sulla base di una serie storica più breve (14 anni, contro i 33 e 22 anni delle altre due serie) e potrebbe quindi risultare meno affidabile. È inoltre da considerare che nessuna delle tre popolazioni è chiusa dal punto di vista biologico, quindi lo scambio di individui tra i tre parchi e il territorio circostante (non quantificato in questo lavoro) potrebbe costituire un ulteriore elemento di incertezza. I

II
Abstract The high-altitude alpine territories are rich in biodiversity and very sensitive to climate change: in fact, they show an increase in temperatures above the global average. The most obvious response of Alpine species to climate change is a shift of populations to higher altitudes. Therefore, species that occupy higher elevations are more vulnerable due to a narrowing in the availability of habitats as the altitude increases. In order to assess the vulnerability of these species in the future, it is crucial to study the demography of alpine wildlife and its dependence on the main environmental variables. In this thesis, population dynamics models were developed for three deer populations (Cervus elaphus) in the Italian Cottian Alps, with the aim of investigating the role of climate change on the demographic dynamics of these species. For this purpose, census data from the 80’s have been collected in three different parks (the Gran Bosco di Salbertrand, Orsiera- Rocciavrè and the Val Troncea) and provided by the Parks Management Authority of the Cottian Alps. The assessment was based on a large set of dynamic models in which the demographic parameters were expressed as a function of density and the main meteorological factors (temperature, precipitation, snow depth). The selection of the best models (in terms of predictive capacity and parsimony) was carried out using two different model selection criteria (the Aikake's Information Criterion and the Bayesian Information Criterion). The selected models incorporate the role of snow depth and cumulative precipitation in influencing the dynamics of the species. These environmental factors, together with population density, interact in determining the demographics of all three populations, despite the presence of some important differences between one population and another. The models calibrated through the reconstruction of the historical trends of the three populations were then used to predict the future demographic dynamics through stochastic simulations (i.e. Monte Carlo simulations). The models were forced by means of synthetic time series of environmental variables corresponding to different possible climate change scenarios (i.e. RCP 4.5 and RCP 8.5 scenarios produced by the IPCC). The results of the future simulations do not show significant risks for two of the three populations (Gran Bosco di Salbertrand and Orsiera-Rocciavrè). For the third (Val Troncea), however, the forecasts obtained with both scenarios indicate a high level of risk. It should be noted, however, that the model relative to the population of the Val Troncea was obtained on the basis of a shorter historical series (14 years, against 33 and 22 years of the other two series) and could therefore be less reliable. It is also to be considered that none of the three populations is closed from the biological point of view, so the exchange of individuals between the three parks and the surrounding territory (not quantified in this work) could represent a further element of uncertainty. III
IV
INDICE SOMMARIO ......................................................................................................................................I ABSTRACT ......................................................................................................................................III NOTAZIONI ................................................................................................................................... VII LISTA DELLE FIGURE ....................................................................................................................... IX INTRODUZIONE ............................................................................................................................... 1 1.1. CONSERVAZIONE E GESTIONE DEGLI UNGULATI SELVATICI .............................................................. 1 1.2. GLI EFFETTI DEI CAMBIAMENTI CLIMATICI ................................................................................... 3 1.2.1 Le Alpi: proprietà e vulnerabilità ....................................................................................4 1.2.2. Effetti sulla fauna alpina .................................................................................................5 1.3. OBIETTIVI E STRUTTURA DELLA TESI .......................................................................................... 6 AREA DI STUDIO E SPECIE D’INTERESSE............................................................................................ 9 2.1. I PARCHI DELLE ALPI COZIE ..................................................................................................... 9 2.1.1. Il Parco Naturale del Gran Bosco di Salbertrand ............................................................10 2.1.2. Il Parco Naturale Orsiera Rocciavrè ................................................................................11 2.1.3. Il Parco Naturale della Val Troncea ................................................................................12 2.2. SPECIE D’INTERESSE – IL CERVO ............................................................................................. 13 2.2.1. Status ..............................................................................................................................13 2.2.2. Distribuzione ecologica ..................................................................................................13 2.2.3. Riproduzione ..................................................................................................................13 2.2.4. Stato di conservazione ...................................................................................................14 DATI DEMOGRAFICI E VARIABILI AMBIENTALI ............................................................................... 15 3.1. DATI DEMOGRAFICI SULLE POPOLAZIONI DI CERVO ..................................................................... 16 3.1.1. Il cervo del Gran Bosco di Salbertrand ...........................................................................16 3.1.2. Il cervo dell’Orsiera Rocciavrè ........................................................................................19 3.2. VARIABILI AMBIENTALI ......................................................................................................... 24 MODELLI DEMOGRAFICI ................................................................................................................ 29 4.1. MODELLI UTILIZZATI ............................................................................................................ 31 4.1.1. Modello esponenziale ....................................................................................................31 4.1.2. Modello di Ricker ...........................................................................................................32 4.1.3. Modello di Beverton-Holt ...............................................................................................32 4.2. SELEZIONE DEI MODELLI ....................................................................................................... 33 4.2.1. Il problema di stima ........................................................................................................34 4.2.2. Scelta del miglior modello ..............................................................................................35 4.2.3. Valutazione dei modelli ..................................................................................................37 RISULTATI E DISCUSSIONE ............................................................................................................. 39 5.1. SIMULAZIONI STORICHE ....................................................................................................... 42 5.1.1. Gran Bosco di Salbertrand ..............................................................................................42 5.1.2. Orsiera-Rocciavrè ...........................................................................................................45 5.1.3. Val Troncea .....................................................................................................................46 5.2. SIMULAZIONI A LUNGO TERMINE ............................................................................................ 48 Osservando la Tabella 5.6, si nota che tutti i modelli hanno residui distribuiti come una normale con media nulla e varianza nota, ad eccezione del modello RT1...............................48 5.2.1. Gran Bosco di Salbertrand ..............................................................................................49 V
5.2.2. Orsiera-Rocciavrè ...........................................................................................................52 5.2.3. Val Troncea .....................................................................................................................55 CONCLUSIONI E SVILUPPI FUTURI .................................................................................................. 57 BIBLIOGRAFIA ............................................................................................................................... 61 RINGRAZIAMENTI .......................................................................................................................... 67 VI
NOTAZIONI Bt tasso di natalità C capacità portante dell’ambiente Dt tasso di mortalità Et tasso di emigrazione It tasso di immigrazione K lunghezza del vettore dei parametri k numero variabili esplicative mj modelli candidati M set di modelli candidati n numero di osservazioni N0 abbondanza iniziale di una popolazione Nt abbondanza di popolazione al tempo t Pt precipitazione cumulata nell’anno t r tasso intrinseco di crescita R2 coefficiente di determinazione R!! coefficiente di determinazione aggiustato St altezza media della neve tra novembre dell’anno t e novembre dell’anno t+1 Tmaxt temperatura massima media nell’anno t Tmint temperatura minima media nell’anno t Tt temperatura media nell’anno t X matrice disegno (n x k) y dati osservati !! stime prodotte da un modello y! valore medio delle osservazioni Simboli greci α costante di crescita β forza della competizione interspecifica β0 parametro costante βi parametro della variabile ambientale i ε vettore degli errori θ vettore dei parametri ! vettore delle stime dei parametri λ massimo tasso di crescita in assenza di competizione VII
Simboli e abbreviazioni AIC Aikake’s Information Criterion AICbest minimo valore dell’AIC AICc Aikake’s Information Criterion del secondo ordine BIC Bayesian Information Criterion BICbest minimo valore del BIC ERA-Interim data-set di rianalisi climatica IFS Integrated Forecasting System IPCC Intergovernmental Panel on Climate Change LS least square MLE maximum likelihood estimation RCP Representative Concentration Pathways SSE sum of square errors VIII
Lista delle Figure Figura 1.1: Cambiamenti osservati in (a) temperatura superficiale media globale, (b) livello medio globale del mare (blu) e satellite (rosso), (c) copertura nevosa dell'emisfero settentrionale nel periodo marzo-aprile e (d) missioni antropogeniche annuali e globali di anidride carbonica (CO2)……………………………………………………………………………………………3 Figura 1.2: Effetti attesi di diversi fattori nel modificare la biodiversità per l'anno 2100. Il riquadro (a) è una media delle stime per ciascun bioma, fatta in relazione al massimo cambiamento; la figura (b) riporta solo gli effetti sul bioma alpino. Adattato da (Sala et al., 2000)……………………………………………………………………………………………………………………….4 Figura 2.1: Inquadramento territoriale dei Parchi delle Alpi Cozie……………………………………….10 Figura 3.1: Censimenti primaverili e al bramito effettuati all’interno del Parco del Gran Bosco di Salbertrand nel periodo 2002-2016……………………………………………………………………..….17 Figura 3.2: Censimenti primaverili e cervi abbattuti nel periodo 1982-2015 all’interno del Parco del Gran Bosco di Salbertrand……………………………………………………………………………17 Figura 3.3: Andamento temporale delle tre specie presenti all’interno del Parco del Gran Bosco di Salbertrand nel periodo 1983-2015……………………………………………………………….18 Figura 3.4: Cervi censiti nel periodo primaverile all’interno del Parco Orsiera-Rocciavrè (1986-2007)………………………………………………………………………………………………………………...19 Figura 3.5: Andamento temporale delle tre specie presenti all’interno del Parco Orsiera- Rocciavrè nel periodo 1997-2007………………………………………………………………………………..20 Figura 3.6: Censimenti al bramito sulla specie cervo del Parco della Val Troncea nel periodo 2002-2015……………………………………………………………………………………………………………………21 Figura 3.7: Andamento temporale delle tre specie presenti all’interno del Parco della Val Troncea nel periodo 2002-2015…………………………………………………………………………………..21 Figura 3.8: Andamento temporale delle tre popolazioni di cervo presenti all’interno delle aree considerate………………………………………………………………………………………………………….22 Figura 3.9: Andamento temporale delle densità delle tre popolazioni nel periodo 2002- 2007…………………………………………………………………………………………………………………………….23 Figura 3.10: Temperatura media annuale nei tre Parchi (1982-2016), elaborazione su dati ERA-Interim…………………………………………………………………………………………………………………25 Figura 3.11: Altezza media della neve (novembre-maggio) nei tre Parchi (1982-2015), elaborazione su dati ERA-Interim………………………………………………………………………………..25 Figura 3.12: Precipitazione cumulata annua nei tre Parchi (1982-2016), elaborazione su dati ERA-Interim…………………………………………………………………………………………………………………25 Figura 3.13: Confronto tra le temperature media annuali ERA-Interim e dati osservativi del Lago Pilone (Parco del Gran Bosco di Salbertrand) nel periodo 1990-2015………………….26 Figura 3.14: Dati sulla popolazione totale del cervo del Parco della Val Troncea e altezza media della neve calcolata come la media tra novembre dell’anno t e maggio dell’anno t+1……………………………………………………………………………………………………………………………….26 Figura 3.15: Proiezioni meteorologiche nell’area dei Parchi delle Alpi Cozie (dati IPCC). (a) Temperatura media annuale; (b) altezza media della neve al suolo tra novembre dell’anno t e maggio dell’anno t+1; (c) precipitazione cumulata annua……………………….28 Figura 4.1: Schema dei fattori principali che influenzano lo stato di una popolazione…………………………………………………………………………………………………………………29 IX
Figura 5.1: Simulazioni dell'abbondanza di popolazione di cervo nel Parco del Gran Bosco di Salbertrand ottenute utilizzando i 6 migliori modelli selezionati………………………………….43 Figura 5.2: Simulazioni sull'abbondanza di popolazione di cervo del Gran Bosco di Salbertrand ottenute utilizzando i 2 migliori modelli selezionati………………………………….44 Figura 5.3: Simulazioni dell'abbondanza di popolazione di cervo nel Parco Orsiera-Rocciavrè ottenute utilizzando i 6 migliori modelli selezionati…………………………………………………….46 Figura 5.4: Simulazioni dell'abbondanza di popolazione di cervo nel Parco della Val Troncea ottenute utilizzando i 2 migliori modelli selezionati…………………………………………………….47 Figura 5.5: Simulazioni a lungo termine dell'abbondanza di popolazione eseguite utilizzando i 6 migliori modelli selezionati per il Parco del Gran Bosco di Salbertrand. Proiezioni ottenute utilizzando i valori medi storici delle variabili ambientali d’interesse…………….49 Figura 5.6: Simulazioni a lungo termine dell'abbondanza di popolazione eseguite utilizzando i 6 migliori modelli selezionati per il Parco del Gran Bosco di Salbertrand. Proiezioni IPCC con scenario di cambiamento climatico RCP 4.5………………………………………………………….50 Figura 5.7: Proiezioni future dell’abbondanza della popolazione del Parco del Gran Bosco di Salbertrand utilizzando due dei migliori modelli e le proiezioni meteorologiche fornite dall’IPCC con scenario di cambiamento climatico RCP 4.5……………………………………………50 Figura 5.8: Simulazioni a lungo termine dell'abbondanza di popolazione eseguite utilizzando i 6 migliori modelli selezionati per il Parco del Gran Bosco di Salbertrand. Proiezioni IPCC con scenario di cambiamento climatico RCP8.5…………………………………………………………..51 Figura 5.9: Proiezioni future sull’abbondanza della popolazione del Parco del Gran Bosco di Salbertrand utilizzando due dei migliori modelli e le proiezioni meteorologiche fornite dall’IPCC con scenario di cambiamento climatico RCP 8.5……………………………………………51 Figura 5.10: Simulazioni a lungo termine dell'abbondanza di popolazione eseguite utilizzando i 6 migliori modelli selezionati per il Parco Orsiera-Rocciavrè. Proiezioni ottenute utilizzando i valori medi storici delle variabili ambientali d’interesse…………………………..52 Figura 5.11: Simulazioni a lungo termine dell'abbondanza di popolazione eseguite utilizzando i 6 migliori modelli selezionati per il Parco Orsiera-Rocciavrè. Proiezioni IPCC con scenario di cambiamento climatico RCP 4.5…………………………………………………………………53 Figura 5.12: Proiezioni future dell’abbondanza della popolazione del Parco Orsiera-Rocciavrè utilizzando due dei migliori modelli e le proiezioni meteorologiche fornite dall’IPCC con scenario di cambiamento climatico RCP 4.5…………………………………………………………………53 Figura 5.13: Simulazioni a lungo termine dell'abbondanza di popolazione eseguite utilizzando i 6 migliori modelli selezionati per il Parco Orsiera-Rocciavrè. Proiezioni IPCC con scenario di cambiamento climatico RCP 8.5…………………………………………………………………54 Figura 5.14: Proiezioni future dell’abbondanza della popolazione del Parco del Gran Bosco di Salbertrand utilizzando due dei migliori modelli e le proiezioni meteorologiche fornite dall’IPCC con scenario di cambiamento climatico RCP 8.5……………………………………………54 X
CAPITOLO 1 Introduzione 1.1. Conservazione e gestione degli ungulati selvatici Negli ultimi decenni l’espansione del popolamento degli ungulati selvatici ha rappresentato uno dei più considerevoli mutamenti del quadro faunistico italiano. Le profonde trasformazioni culturali, socio-economiche e normative avvenute a partire dalla metà del secolo scorso hanno determinato condizioni favorevoli a una progressiva riconquista degli antichi areali e oggi gli ungulati hanno ripopolato una considerevole parte del territorio nazionale, dopo che per almeno tre secoli l’azione dell’uomo aveva provocato la loro sostanziale scomparsa. Essi sono tornati dunque a svolgere un importante ruolo strutturale e funzionale nelle biocenosi italiane, con indubbie ricadute positive anche per la conservazione di altre specie e in particolare dei grandi carnivori (Toso, 2013). Grazie alla diffusione degli ungulati avvenuta negli ultimi tre decenni, sono state sviluppate modalità di gestione basate sul principio della sostenibilità dell’utilizzo di questa risorsa nelle sue diverse forme. Malgrado questo processo presenti ancora lacune, discontinuità e una diffusione disomogenea in ambito nazionale ed europeo, esso ha rappresentato un chiaro elemento di crescita culturale, con evidenti ripercussioni positive che non riguardano solo un uso ecologicamente compatibile delle popolazioni degli ungulati, ma che investono l’approccio generale alla conservazione della fauna selvatica. Gli ungulati, che in Italia comprendono cervo, capriolo, camoscio, muflone, stambecco, cinghiale e daino, sono di fondamentale importanza per il loro ruolo dominante nella struttura e nelle dinamiche ecologiche degli ecosistemi naturali (Putman, 2004) e vengono considerati sia come risorsa, estetica o economica, che come fonte di problemi derivanti dall’impatto che la loro presenza determina sull’agricoltura, i boschi e la sicurezza stradale. Molte specie si nutrono, infatti, in maniera molto selettiva e, di conseguenza, possono 1
modificare la composizione e l'abbondanza delle diverse componenti della vegetazione, così come la loro struttura fisica (Putman, 1986, 2004). A causa dell’impatto sulla componente vegetale, questi grandi erbivori possono condizionare fortemente le comunità animali e in particolare gli insetti, gli uccelli e la piccola fauna, con effetti a catena sulla popolazione di predatori, i quali dipendono fortemente dalla disponibilità di prede come gli ungulati (Putman, 1986,1994; Putman et al., 1989; Petty and Avery, 1990; Fuller, 2001; Flowerdew and Ellwood, 2001; Suominem and Danell, 2006). Gli ungulati non sono però importanti soltanto perché prede naturali del lupo e della lince, ma anche perché rappresentano un'alternativa alla predazione sugli animali domestici (Meriggi & Lovari, 1996), una delle principali cause degli attuali conflitti tra i grandi carnivori e l’uomo (Gazzola et al., 2008). E’ riconosciuto inoltre che la presenza stabile di popolazioni di grandi erbivori in territori occupati dall’uomo può entrare in conflitto con gli obiettivi che quest’ultimo ha sull’uso del suolo e le sue risorse (Motta, 1998; Putman & Moore, 1998; Ammer, 1996). In passato il principale danno causato dagli ungulati avveniva in agricoltura (Putman & Kjellander, 2003), oggi, invece, i boschi e le foreste sembrano subire l’impatto più rilevante (Mayer & Charlesworth, 1991; Welch et al., 1991, 1992; van Hees et al., 1996; Reimoser, 2000). A causa del brucamento e dell’asportazione della corteccia dagli alberi, in aree con un’alta densità di ungulati si può verificare una riduzione delle possibilità di rigenerazione naturale di ampie zone boschive (Remoser & Gossow, 1996; van Hees et al., 1996; Vera et al., 2006), con implicazioni negative sull’intero ecosistema. Storicamente, il sovrasfruttamento, la predazione, le malattie e i cambiamenti nell'uso del suolo hanno ridotto il numero di molte specie di ungulati (Teer, 1996; Danz, 1997). Tuttavia, in diverse zone del mondo alcune specie hanno beneficiato dei cambiamenti climatici e delle trasformazioni nell’uso del suolo, accrescendo il loro numero e la loro distribuzione su vasti territori. La riduzione del prelievo da parte dell’uomo e la mancanza di predatori ha portato dunque a una situazione tale da dover assicurare che il loro numero non incida su altri obiettivi di uso del suolo, compresi l'agricoltura e la silvicoltura, e non influisca negativamente sugli ecosistemi in generale (Cederlund et al., 1998; McShea, Underwood & Rappole, 1997). La conservazione dei grandi erbivori selvatici, intesa nel senso più ampio del termine, includendo anche la gestione attiva attraverso il prelievo, rappresenta oggi una delle attività più rilevanti per gli organismi gestori e le loro scelte di carattere normativo, programmatico ed operativo devono essere fondate su solide basi scientifiche e tecniche. La gestione di queste popolazioni di ungulati deve quindi basarsi su un'approfondita conoscenza scientifica dell'ecologia e delle dinamiche demografiche di queste specie e dei sistemi più ampi di cui fanno parte. 2
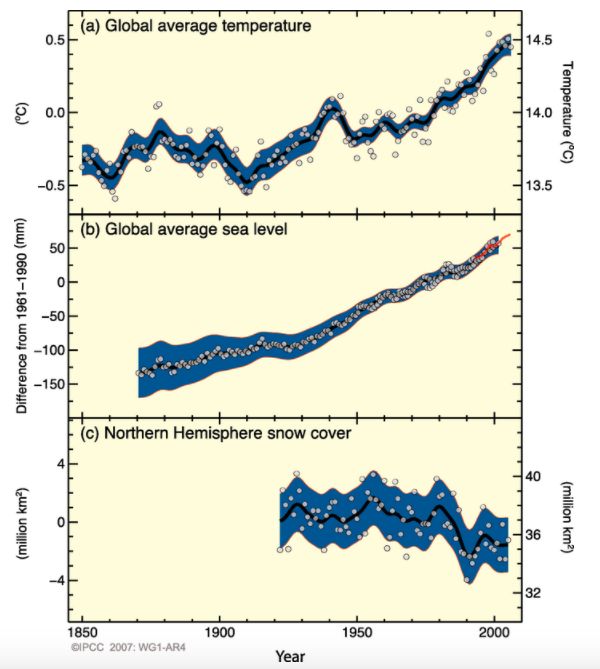
1.2. Gli effetti dei cambiamenti climatici I cambiamenti climatici si riferiscono a un mutamento nello stato del clima che può essere identificato (ad esempio utilizzando test statistici) modificando la media e/o la variabilità delle sue proprietà e che persiste per un periodo prolungato, tipicamente lungo decenni o più. Si riferisce a qualsiasi cambiamento climatico nel corso del tempo, causato dalla variabilità naturale o come risultato dell'attività umana (IPCC, 2007). Il riscaldamento del sistema climatico è inequivocabile e, a partire dagli anni '50, molti dei cambiamenti osservati sono senza precedenti. L'atmosfera e l'oceano si sono riscaldati, le quantità di neve e ghiaccio sono diminuite e il livello del mare è aumentato (vedi Figura 1.1). Le emissioni antropogeniche di gas serra sono aumentate dall'era preindustriale, trainate in gran parte dalla crescita economica e demografica, e ora sono più alte che mai. Ciò ha portato a concentrazioni atmosferiche di anidride carbonica, metano e protossido di azoto che non hanno precedenti negli ultimi 800.000 anni. I loro effetti, insieme a quelli di altre forzanti antropogeniche, sono stati rilevati in tutto il sistema climatico ed è estremamente probabile che siano stati la causa principale del riscaldamento osservato dalla metà del XX secolo (IPCC (AR5), 2014). Tale cambiamento sta avendo effetti significativamente impattanti sui sistemi naturali. In molte regioni, il cambiamento delle precipitazioni o lo scioglimento di neve e ghiaccio stanno alterando i sistemi idrologici e influenzando le risorse idriche in termini di quantità e qualità. Molte specie terrestri, di acqua dolce e marine hanno modificato le preferenze degli habitat, le attività stagionali, le abbondanze e le interazioni tra le specie, in risposta ai cambiamenti climatici in corso. (d) Figura 1.1: Cambiamenti osservati in (a) temperatura superficiale media globale, (b) livello medio globale del mare (blu) e satellite (rosso), (c) copertura nevosa dell'emisfero settentrionale nel periodo marzo-aprile e (d) missioni antropogeniche annuali e globali di anidride carbonica (CO2). Tutte le modifiche sono relative alle medie corrispondenti per il periodo 1961-1990. Le curve arrotondate rappresentano valori medi decennali mentre i cerchi mostrano valori annuali. (Fonte IPCC, 2007, WG1-AR4). 3
1.2.1 Le Alpi: proprietà e vulnerabilità Le regioni montane rappresentano circa il 20-24% della superficie terrestre e sono caratterizzate da diversi tipi di clima e, anche se la ricchezza di specie diminuisce con l'altitudine, l’ambiente montano è caratterizzato da molti ecosistemi diversi e presenta quindi una delle più grandi ricchezze di specie a livello globale (Väre et al., 2003; Moser et al., 2005; Spehn & Körner, 2005). Tra le regioni montane, le Alpi sono di particolare interesse poiché ospitano una grande quantità di specie endemiche e sono perciò considerate tra le regioni più importanti per la conservazione della biodiversità in Europa (Theurillat & Guisan, 2001; Korner & Spehn, 2002). Tuttavia le regioni montane, e in particolare quelle europee, sono molto vulnerabili ai cambiamenti climatici e ai disturbi antropogenici (Fischlin et al., 2007). In queste regioni, infatti, si possono riscontrare aumenti di temperatura più elevati rispetto alle regioni circostanti. Nelle Alpi la temperatura sta aumentando a un ritmo circa doppio rispetto alla media globale osservata nell’ultimo secolo (Auer et al., 2007). L’attuale configurazione dell'ambiente alpino è il risultato dell’interazione tra condizioni naturali e secoli di cambiamenti dovuti all’azione dell’uomo. Ad esempio, il recente abbandono dei pascoli alpini, insieme ai cambiamenti climatici, ha causato un'espansione dei boschi e uno spostamento verso l'alto della linea degli alberi (Schweiger et al., 2012), con conseguenti ripercussioni negative sulla biodiversità locale (Dirnböck et al., 2003; Laiolo et al., 2004; Martin & Possingham, 2005). Uno studio di Sala et al. (2000) ha dimostrato che, mentre a livello mondiale il principale driver della perdita di biodiversità prevista per l'anno 2100 è il cambiamento di uso del suolo (vedi Figura 1.2 (a)), sul bioma alpino è previsto un maggiore impatto dovuto al cambiamento climatico, come riportato nella Figura 1.2 (b). Gli ecosistemi alpini sono caratterizzati, infatti, da basse temperature e ospitano organismi altamente specializzati che vivono vicino al limite delle loro tolleranze fisiologiche (Mignatti, 2014), e per questo motivo dovrebbero mostrare gli effetti del cambiamento climatico in anticipo e in modo più evidente rispetto ad altri ecosistemi (IPCC, 2007a; Pauli et al., 2007; Pickering et al., 2008). 1,2 0,6 Relative effect of drivers 1 Relative biodiversity change 0,5 0,8 0,6 0,4 0,4 0,3 0,2 0 0,2 0,1 0 1 2 3 4 5 (a) A livello mondiale (b) Bioma alpino Figura 1.2: Effetti attesi di diversi fattori nel modificare la biodiversità per l'anno 2100. Il riquadro (a) è una media delle stime per ciascun bioma, fatta in relazione al massimo cambiamento; la figura (b) riporta solo gli effetti sul bioma alpino. Adattato da Sala et al. (2000). 4
1.2.2. Effetti sulla fauna alpina Le specie che vivono in ambienti montani possono essere particolarmente sensibili ai cambiamenti climatici (Post et al. 1999) e possono andare incontro a un incremento del rischio di estinzione (Derocher & Stirling 1995, 1998). Gli effetti registrati e attesi del riscaldamento globale sulla fauna sono principalmente legati a cambiamenti nella fisiologia, fenologia e distribuzione spaziale. Teoricamente, la risposta della fauna alle trasformazioni ambientali può essere più veloce rispetto alla risposta della vegetazione, perché le specie in costante movimento non hanno bisogno di aspettare fino alla generazione successiva per cambiare posizione o distribuzione sul territorio. Questa caratteristica aumenta, quindi, la plasticità della nicchia ecologica degli animali e li rende più efficienti nel rispondere agli eventi estremi. Tra i mammiferi, i maschi dello stambecco alpino hanno mostrato, nel Parco Nazionale Svizzero, un aumento dell'altitudine estiva di circa 250 metri in diciannove anni (Herfindal et al., 2012), aumentando così la sovrapposizione tra i territori occupati dai maschi e dalle femmine. In tale situazione è probabile che aumenti la competizione intraspecifica all’interno della specie stessa e si abbia, di conseguenza, una riduzione della sopravvivenza o della fertilità. Sulla stessa specie, lo stambecco alpino, sono stati condotti molti studi per indagare e approfondire il ruolo del clima sulle dinamiche demografiche della popolazione del Parco Nazionale del Gran Paradiso (GPNP), in Italia (ad esempio Jacobson et al., 2004; Lima & Berryman, 2006). Il risultato di questi studi ha identificato come principale effetto una diminuzione della sopravvivenza invernale associata ad una elevata profondità media della neve durante l’inverno. Le regioni alpine di alta quota sono caratterizzate da una grande importanza ecologica e una forte sensibilità alle condizioni climatiche. Il cambiamento climatico sta interessando il biota di montagna in diversi modi: in modo diretto, ad esempio attraverso l'aumento delle temperature e la variazione del regime di precipitazione e, in maniera indiretta, attraverso la variazione delle componenti abiotiche dell'ambiente (ad esempio la riduzione dei ghiacciai). Le risposte ecologiche a queste pressioni dipendono dalle caratteristiche specifiche di ciascuna specie e le risposte di ogni singola specie possono portare a una variazione nell'interazione tra diverse specie allo stesso livello o a diversi livelli trofici, e a una variazione più generale delle comunità ecologiche. Generalmente si ritiene che il rigido clima invernale influenzi la sopravvivenza degli individui giovani attraverso una combinazione di maggiori costi di termoregolazione e una diminuzione della disponibilità di foraggio a causa della presenza di neve al suolo (Forchhammer et al., 1998b; Loison & Langvatn, 1998; Portier et al., 1998). E’ quindi essenziale testare la capacità dei modelli di dinamica di popolazione, non soltanto di spiegare gli andamenti demografici storici, ma anche di prevedere i cambiamenti futuri (Forchhammer et al. 1998a), basati sulle proiezioni climatiche. 5
1.3. Obiettivi e struttura della tesi La comprensione di quali fattori determinano l'aumento, la diminuzione o la stabilità delle popolazioni di ungulati selvatici è di fondamentale importanza per acquisire informazioni utili alla gestione delle stesse. Molti studi ecologici a lungo termine sulla dinamica della popolazione di questi grandi erbivori hanno fornito una comprensione dettagliata degli effetti di alcuni fattori nel determinare la dimensione e la composizione di una popolazione (Saether 1997; Gaillard, Festa-Bianchet & Yoccoz 1998; Gaillard et al., 2000). Questi studi si sono concentrati sulle relazioni tra densità di popolazione, condizioni meteorologiche e tassi di sopravvivenza individuale di diverse classi di sesso/età (Gaillard, Festa-Bianchet & Yoccoz 1998, Gaillard et al., 2000) e hanno evidenziato che gli impatti di entrambi gli effetti, sia dipendenti sia indipendenti da densità, sono sostanzialmente correlati con il sesso/struttura di una popolazione. Ciò è dovuto al fatto che la sopravvivenza di individui di diverso sesso/età non è ugualmente influenzata dall'abbondanza di risorse e dai fattori meteorologici. In generale, gli adulti sono relativamente insensibili alla densità e agli effetti meteorologici, mentre i giovani (e forse gli individui senescenti) sono molto suscettibili a entrambi (Gaillard, Festa-Bianchet e Yoccoz 1998; Gaillard et al., 2000; Coulson et al., 2001). La dinamica delle popolazioni di ungulati è pertanto fortemente determinata sia dalla dipendenza da densità, sia dai driver ambientali (Forchhammer et al., 2002; Lande, 1993; Post et al., 1997; Sæ ther & Saether, 1997), che possono operare in modo sinergico (Gaillard & Yoccoz, 2003). L’obiettivo di questa tesi è lo studio della dinamica delle popolazioni di ungulati alpini in un contesto in cui le variabili ambientali, in particolare quelle climatiche, possono influenzare la dinamica temporale di queste specie e condizionarne le modalità di gestione future. Nello studio è stata sfruttata l’insolita opportunità di avere a disposizione i dati delle serie temporali di tre popolazioni di cervo per testare la capacità di una serie di modelli di prevederne la dinamica demografica. Per riprodurre le dinamiche di popolazione e fare previsioni nel prossimo futuro, sono stati proposti diversi modelli. Un approccio semplice ma robusto è stato proposto in passato da Jacobson et al. (2004), che ha esplorato diverse possibili relazioni tra la crescita della popolazione totale di stambecchi nel Parco Nazionale del Gran Paradiso (GPNP) e la profondità della neve nel periodo invernale. Nel presente studio, i modelli proposti da Jacobson et al. (2004) sono stati modificati nella loro struttura e ampliati grazie all’introduzione di altre variabili ambientali. Innanzitutto sono stati stimati gli effetti indipendenti delle variabili meteorologiche e della densità sui cambiamenti annuali delle dimensioni delle tre popolazioni di cervo che occupano i territori dei Parchi delle Alpi Cozie, in Italia. Una volta dimostrato che queste variabili influenzano la dinamica di popolazione, è stato sviluppato un modello che tiene conto contemporaneamente di tutti i fattori. Successivamente, è stato valutato il potere predittivo a lungo termine di questo modello. La tesi si articola nei seguenti capitoli: • il capitolo 1 spiega il contesto in cui è stato svolto lo studio, specificando le problematiche relative alla presenza di ungulati in ambiente alpino; 6
• il capitolo 2 descrive l’area di studio su cui sono state svolte le indagini, con una descrizione generale della specie in esame; • il capitolo 3 riporta le modalità di acquisizione e di elaborazione dei dati demografici e meteorologici utilizzati nello studio; • il capitolo 4 esamina le dinamiche temporali e gli effetti non lineari delle variabili climatiche sui parametri demografici e la loro relazione con la densità di popolazione. Particolare attenzione è stata posta sull’identificazione delle strutture del modello e delle variabili ambientali più influenti. Questa analisi, detta model selection, include la formulazione delle ipotesi ecologiche, la loro traduzione in modelli matematici, l’identificazione del modello (o calibrazione dei parametri) e la scelta del miglior modello (o dei migliori modelli); • il capitolo 5 mostra i risultati delle analisi, con particolare attenzione agli scenari di cambiamento attesi per il futuro; • Il capitolo 6 contiene le conclusioni a cui si è giunti a seguito delle analisi condotte nell’ambito dello studio e si fa riferimento ai possibili sviluppi futuri. 7
8
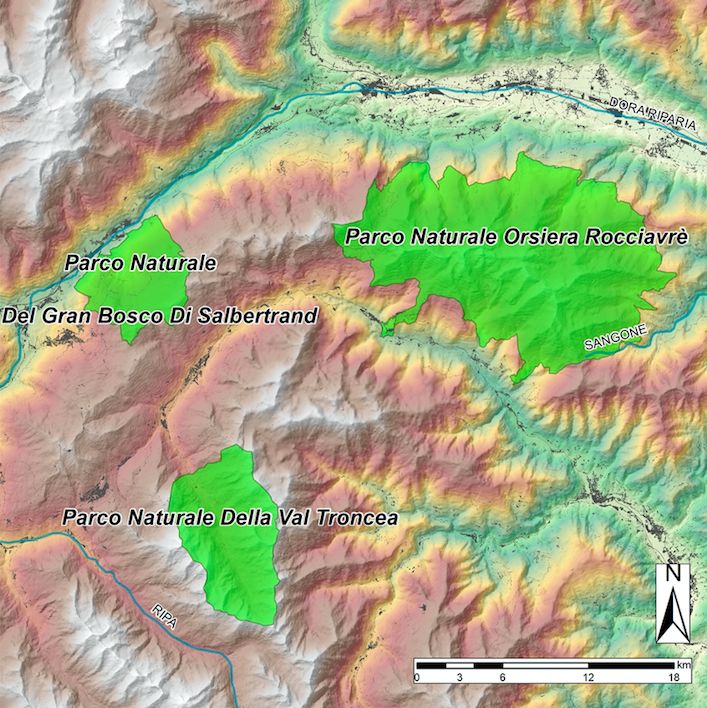
CAPITOLO 2 Area di studio e specie d’interesse 2.1. I Parchi delle Alpi Cozie L’area esaminata dal presente studio è situata nel territorio alpino della Regione Piemonte, nel nord-ovest dell’Italia (vedi Figura 2.1), e comprende tre dei quattro Parchi Naturali amministrati dall’Ente di Gestione dei Parchi delle Alpi Cozie. Istituiti nel 1980, questi Parchi si estendono per una superficie complessiva di oltre 18.000 ettari e coprono un territorio vasto che dalla pianura sale fino a superare i 3.500 m s.l.m.. Si tratta di un parco d'acqua (Parco naturale dei Laghi di Avigliana) e di tre parchi naturali di montagna (Parco naturale della Val Troncea, Parco naturale dell'Orsiera Rocciavré e Parco naturale del Gran Bosco di Salbertrand). I Parchi naturali Val Troncea, Orsiera Rocciavré e Gran Bosco di Salbertrand tutelano ambienti tipicamente alpini, ricchi di fauna selvatica, e rappresentano, dunque, territori adatti allo studio della dinamica delle popolazioni di ungulati selvatici che occupano queste aree protette. Al contrario, all’interno del Parco naturale dei Laghi di Avigliana non è presente alcuna popolazione di ungulati. Quest’ultimo Parco si occupa principalmente della tutela delle zone umide e del ripristino delle condizioni idrobiologiche dei laghi e, per questo motivo, il territorio di questa area protetta non è stato oggetto d’indagine del nostro studio. Dall’1 gennaio 2012 i Parchi naturali dei Laghi di Avigliana, Orsiera Rocciavrè, Gran Bosco di Salbertrand e Val Troncea sono stati unificati nell'Ente di Gestione delle Aree Protette delle Alpi Cozie. Grazie a questo accorpamento, le modalità di monitoraggio degli ungulati dei tre Parchi sono state standardizzate, con ripercussioni positive sia dal punto di vista della gestione di tali popolazioni, sia per quanto riguarda la conservazione delle stesse. 9
FRANCIA ITALIA Figura 2.1: Inquadramento territoriale dei Parchi delle Alpi Cozie. 2.1.1. Il Parco Naturale del Gran Bosco di Salbertrand L'Area Protetta del Parco naturale del Gran Bosco di Salbertrand (45°03′58.53″N 6°55′21.64″E) occupa una superficie di 3.774,74 ettari e si estende sulla destra orografica della Val di Susa (Alpi Cozie Settentrionali), dai 1.000 m fino a raggiungere i 2.700 m s.l.m.. Il Parco del Gran Bosco è stato istituito al fine di tutelare un nucleo forestale tra i più interessanti delle Alpi piemontesi, sia per la varietà della composizione dendrologica, sia per la sua produttività, sia ancora per lo stato di conservazione che sottintende un notevole valore paesaggistico e naturalistico per quanto riguarda la vegetazione. Il Gran Bosco presenta una zonazione altitudinale completa e tipica di ambienti forestali alpini. La fauna è particolarmente ricca: conta, infatti, la presenza di quattro specie di ungulati (camoscio, cinghiale, cervo e capriolo). Dal 1997, anno in cui per la prima volta è stata accertata la presenza di un branco di lupi all’interno dell’area del Gran Bosco, si sono susseguiti numerosi progetti di studio e monitoraggio finanziati dalla Comunità Europea, che oggi proseguono con il personale del Parco, sotto la direzione dei ricercatori del Centro Grandi Carnivori del Parco Alpi Marittime. All’interno del Parco il problema di maggiore rilievo non è dato dall'impatto antropico, che potrà però diventare più marcato nel prossimo futuro, ma dal carico di ungulati selvatici, soprattutto dei cervi (Perco, 1995), la cui massiccia presenza crea problemi di conservazione del soprassuolo forestale. Infatti, per sopperire alle esigenze alimentari dei mesi invernali, i cervi si nutrono di giovani piantine di abete, mentre sulle piante adulte, in particolare sulle latifoglie, sfregando il proprio trofeo, creano delle scortecciature giungendo anche a causare lesioni a livello cellulare. L’elevata presenza di cervi, di introduzione artificiale, in un ambiente antropizzato e privo, fino a pochi anni fa, di predatori, costituisce una minaccia per l’ecosistema, a causa dell'eccessiva quota di produzione primaria consumata. 10
Le conseguenze, anche a breve e medio termine, del persistere dell'attuale carico di ungulati all'interno del Parco possono risultare di estrema gravità. Anche nelle zone che attualmente sono meno danneggiate si prevede nei prossimi anni, in seguito ai danneggiamenti da ungulati, una riduzione di accrescimento, una perdita di qualità delle produzioni legnose e la perdita di numerosi giovani alberi, che potrebbero contribuire al miglioramento del patrimonio genetico del bosco. Inoltre la selettività delle preferenze alimentari dei cervidi comporterà verosimilmente sul medio-lungo periodo una variazione della composizione floristica dello strato arboreo a tutto vantaggio delle specie meno appetite che però non sono sempre quelle cui tende la successione naturale (Quaglino & Motta, 1987). Tra gli altri danni all'ecosistema legati alla presenza del cervo e accertati nelle indagini condotte sul territorio in esame, occorre sottolineare anche una sensibile diminuzione della nidificazione del gallo forcello (Tetrao tetrix L.), dovuta alla distruzione di buona parte dello stato arbustivo necessario alla vita di questa specie e comunque al disturbo ad essa arrecato durante alcune fasi delicate del ciclo biologico quali il canto primaverile, la nidificazione e l'allevamento della prole. I danni rilevati sul patrimonio forestale e ambientale del Parco Naturale del Gran Bosco di Salbertrand risultano quindi decisamente superiori al limite sopportabile dell'ambiente. Sulla base di quanto affermato risulta evidente che ogni azione intesa al miglioramento del patrimonio forestale e ambientale del Gran Bosco (ed oggi dell'intera Valle di Susa) trova nel sovraccarico di ungulati forti ostacoli alla sua applicazione. Da più anni il Parco effettua piani di abbattimento nei confronti del cervo, ai fini di ridurre i danni all'ecosistema, mediante una diminuzione della sua consistenza in foresta. Tuttavia i risultati non hanno portato sinora alla soluzione del problema dei danni, bensì a una diminuzione della presenza del cervo, tale però da non sortire effetti benefici per la vegetazione, principalmente perché analoghi provvedimenti sono stati adottati tardivamente all'esterno dell'area protetta (Perco, 1995). Nell'ottica generale del Parco bisognerà dunque trovare un equilibrio tra la gestione del bosco e la presenza dei cervi nel territorio, evitando la regressione e la scomparsa di formazioni boschive non riscontrabili in altri settori del Piemonte e di estremo interesse naturalistico e scientifico in generale. 2.1.2. Il Parco Naturale Orsiera Rocciavrè Il Parco Naturale Orsiera Rocciavrè (45°03ʹ03.57″N 7°12ʹ43.77″E), i cui confini si trovano mediamente a una quota altimetrica di 1.400 m, si estende per 10.928 ettari nelle Alpi Cozie Settentrionali, su territori di pertinenza delle Valli Chisone, Susa e Sangone. Nel Parco sono presenti sei specie di ungulati selvatici: stambecco, camoscio, capriolo, cervo, muflone e cinghiale. Fra tutte solamente il camoscio è sempre stato presente nell'area protetta e nei territori limitrofi, mentre le altre specie sono state oggetto di immissioni artificiali. Cervo e capriolo devono la loro attuale presenza all'espansione demografica dei nuclei costituiti con le operazioni di reintroduzione effettuate nel periodo 1962-1965 nel Gran Bosco di Salbertrand. L’ambiente preferito da questi cervidi è rappresentato da spazi aperti alternati a bosco, ma si adattano molto bene anche all'alta montagna, dove prediligono i pascoli al limite della 11
Puoi anche leggere

















































