CITOLOGIA 10 - Reticolo endoplasmatico, lisosomi, perossisomi, inclusioni cellulari
←
→
Trascrizione del contenuto della pagina
Se il tuo browser non visualizza correttamente la pagina, ti preghiamo di leggere il contenuto della pagina quaggiù
CITOLOGIA 10 - Reticolo endoplasmatico, lisosomi, perossisomi, inclusioni cellulari
Reticolo endoplasmatico
Il reticolo endoplasmatico (RE) è un organulo citoplasmatico, costituito da una rete intrecciata di
compartimenti (detti cisterne).
Le cisterne sono formate da un doppio strato di fosfolipidi, che rende la membrana del RE simile a quella
citoplasmatica; tuttavia, essa è più sottile e presenta catene di oligosaccaridi che sporgono anche
all’interno delle cisterne (e non solo all’esterno, come nel caso della membrana cellulare).
La funzione più importante del RE è quella di modificare alcune sostanze prodotte dalla cellula.
Ci sono 2 tipi di RE: rugoso (RER) e liscio (REL – o SER in inglese).
Reticolo endoplasmatico rugoso (RER)
Il RER deve il suo attributo “rugoso” alla presenza
di ribosomi attaccati sul lato esterno della
membrana che gli conferiscono un aspetto ruvido
(simile ad una carta vetrata vista al microscopio).
La membrana esterna del RER si continua con la
membrana esterna del nucleo. Anche essa
presenta ribosomi sulla superficie esterna.
Le cisterne del RER hanno la caratteristica forma
appiattita e sono molto sviluppate ed abbondanti
nelle cellule dove c’è un elevato metabolismo
proteico (es. cellule del fegato e dell’intestino).
La funzione principale del RER è quella di
rimaneggiare le proteine sintetizzate dai ribosomi
presenti sulla sua superficie esterna. In particolare, le funzioni principali sono la N-Glicosilazione e il folding
proteico.
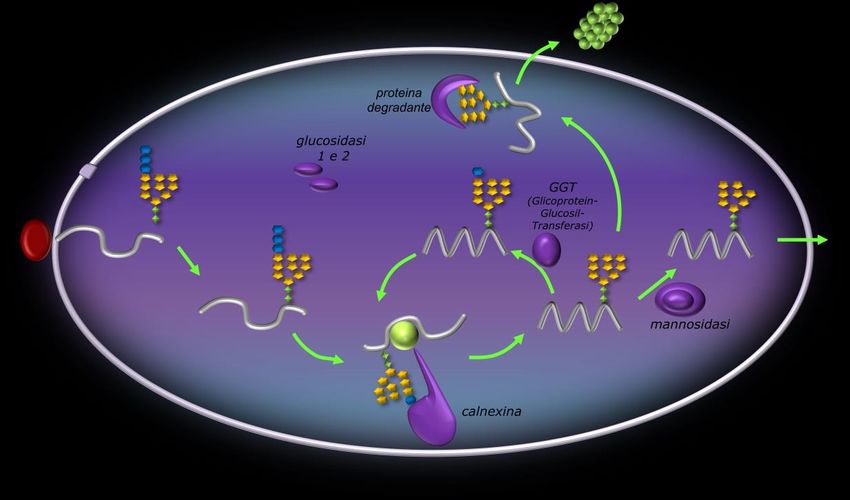
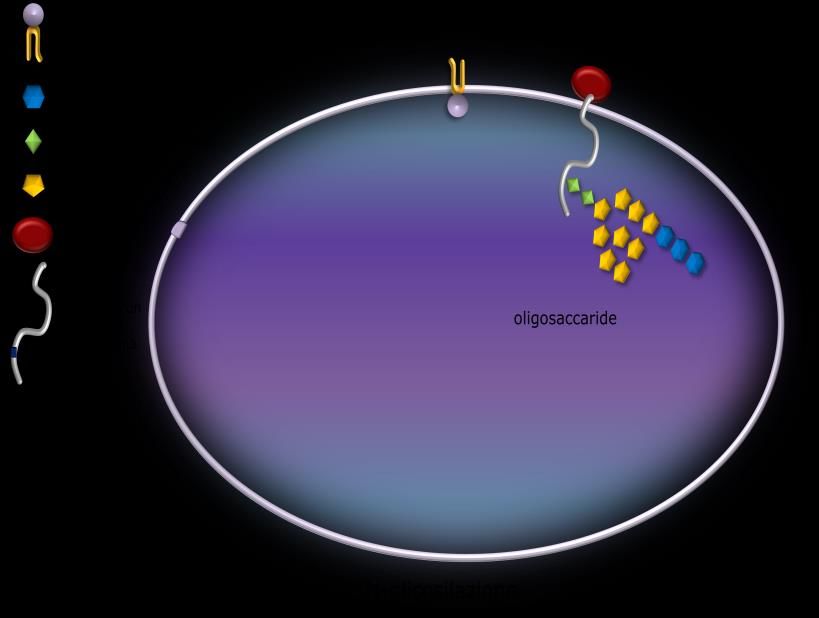
N-glicosilazione
Consiste nell’aggiunta di un
oligosaccaride (formato da 14 residui
zuccherini) alla proteina sintetizzata dai
ribosomi presenti sul RER.
L’oligosaccaride si forma sul dolicolo, un
fosfolipide particolare che si trova in
prossimità del canale d’ingresso
attraverso il quale la proteina
sintetizzata dal ribosoma entra nel RER.
Successivamente, intervengono degli
enzimi (glicosidasi) che eliminano 3
molecole di glucosio ed 1 di mannosio
dall’oligosaccaride.
Infine, quando entra una proteina
appena sintetizzata dal ribosoma,
l’oligosaccaride viene trasferito sul gruppo amminico (NH2) dell’amminoacido asparagina.
La presenza dell’albero oligosaccaridico è fondamentale, perché:
permette alle proteine di essere più solubili
le rende meno attaccabili da parte delle proteasi
1
CITOLOGIA 10 - Reticolo endoplasmatico, lisosomi, perossisomi, inclusioni cellulari
conferisce loro un corretto ripiegamento (folding delle proteine).
Folding proteico
Una proteina, per funzionare correttamente, non deve solo avere la giusta sequenza di amminoacidi, ma
deve assumere anche una corretta conformazione spaziale.
Il folding proteico avviene attraverso diverse fasi:
1. La proteina appena prodotta dal ribosoma ed N-glicosilata viene sottoposta all’azione di glicosidasi
di tipo I e II, che sottraggono all’oligosaccaride 2 molecole di glucosio.
2. A questo punto, interviene una proteina, la calnexina, che si attacca da una parte con il glucosio
rimasto sull’oligosaccaride e dall’altra sulla proteina. La calnexina riesce a fare un primo
ripiegamento della proteina, eliminando l’ultima molecola di glucosio rimasta.
3. La proteina appena ripiegata può andare incontro a due destini:
1. Se il primo ripiegamento della calnexina è sufficiente, la proteina esce dal RER. Prima di
uscire perde, però, una molecola di mannosio dall’oligosaccaride (ad opera dell’enzima
mannosidasi).
2. Se la proteina necessita di più ripiegamenti, allora interviene un’altra proteina, la GGT
(Glicoprotein-Glucosil-Transferasi) che aggiunge una molecola di glucosio all’albero. In
questo modo, la proteina si può legare nuovamente alla calnexina e subire un ulteriore
ripiegamento. Questo percorso ciclico continua fino a quando la proteina non ha subito
tutti i ripiegamenti di cui ha bisogno.
4. A questo punto, le proteine saranno tutte ripiegate, ma non tutte lo saranno correttamente. Le
proteine misfolding (cioè ripiegate male) vengono riconosciute da un’altra proteina, detta proteina
degradante, la quale porta la proteina fuori dal RER verso il proteosoma, un complesso proteico che
distrugge la proteina difettosa.
Reticolo endoplasmatico liscio (REL)
A differenza del RER, il REL è privo di ribosomi sulla sua superficie (di qui “liscio”); le cisterne da cui è
composto hanno una forma tubulare, che lo rende facilmente distinguibile dal RER.
2
CITOLOGIA 10 - Reticolo endoplasmatico, lisosomi, perossisomi, inclusioni cellulari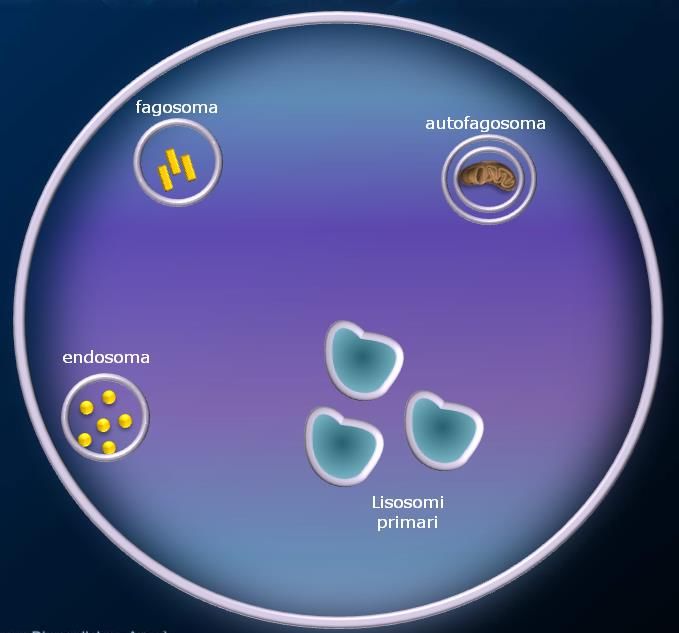
Il REL ha diverse funzioni:
Sintetizzare lipidi (i fosfolipidi che servono per le membrane cellulari, il colesterolo e gli steroidi e i
glicolipidi).
Detossificare da sostanze altrimenti dannose per l’organismo (come l’etanolo contenuto nelle
bevande alcoliche, alcuni farmaci, etc). Questa funzione viene espletata da ossidasi, tra cui i
citocromi P450.
Costituire una riserva di ioni calcio (Ca2+), utili in alcune cellule (come quelle muscolari).
È coinvolto nel metabolismo del glicogeno. Il glicogeno viene conservato, come riserva energetica,
nel fegato e nei muscoli. Per fornire energia, esso deve essere “spezzettato” in tante molecole di
glucosio. Questo processo avviene grazie al REL.
Infatti, il glicogeno viene convertito dapprima in tante molecole di glucosio-6-fosfato (glucosio + P).
In questa forma, però, il glucosio non può uscire dalla cellula. Grazie ad un enzima, la glucosio-6-
fosfatasi, presente solo sulle membrane del REL, il gruppo P viene rimosso e il glucosio può uscire
dalla cellula.
Lisosomi
I lisosomi sono organuli cellulari deputati alla digestione di materiale extra o intracellulare. Si possono
definire “l’apparato digerente della cellula”.
Si tratta di piccole vescicole (tra 25-50 nm e 1 μm), che si formano per gemmazione dalle membrane del
Golgi. I lisosomi contengono una grande varietà di enzimi, chiamati idrolasi acide, capaci di digerire
praticamente tutti i tipi di molecole biologiche (proteine, lipidi, acidi nucleici, glucidi). Questi enzimi sono
prodotti nel RER e rielaborate dal Golgi e funzionano a pH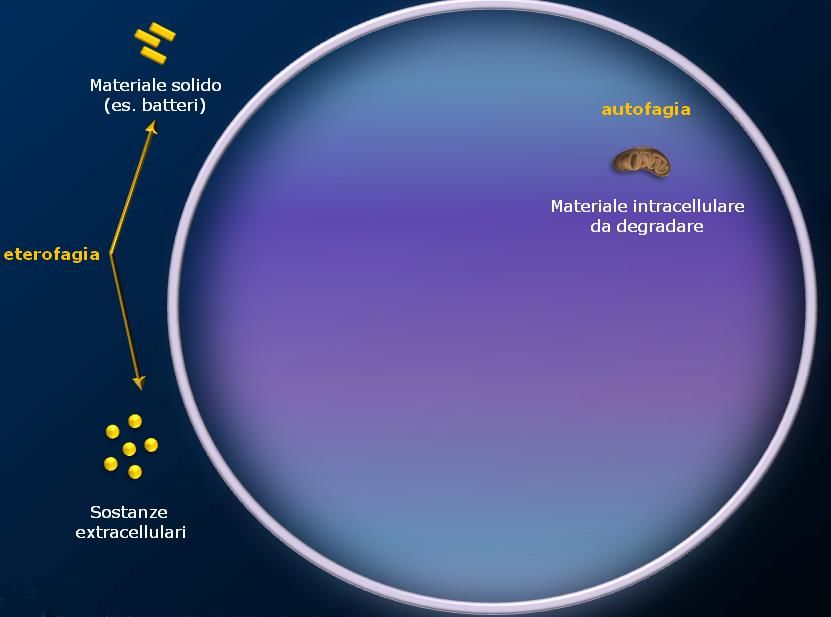
Le cellule che contengono più lisosomi sono i macrofagi e i granulociti, cellule deputate alla difesa del
nostro organismo.
Difetti nella composizione delle idrolasi lisosomiali possono portare a gravi malattie; tra queste, spicca la
malattia di Gaucher, dovuta alla deficienza dell’enzima glucocerebrosidasi, che comporta gravi danni al
fegato, alla milza e alle ossa.
Perossisomi
Sono simili ai lisosomi, anche nelle dimensioni (0,5 μm circa), ma hanno enzimi diversi. Sono così chiamati
perché, grazie agli enzimi perossidasi, riescono a convertire l’acqua (H2O) in acqua ossigenata (H2O2).
Oltre alle perossidasi, i perossisomi contengono altri enzimi, tra cui le catalasi e le ossalasi.
I perossisomi non vengono prodotti nel Golgi (come i lisosomi) ma si riproducono per divisione.
I perossisomi hanno la funzione di degradare gli acidi grassi, l’alcol ed altri componenti dannosi.
Per questo, sono maggiormente presenti nelle cellule del fegato e dei reni, due importanti filtri del nostro
organismo.
Al microscopio elettronico, i perossisomi si differenziano dai lisosomi per l’aspetto delle vescicole.
Infatti, rispetto ai lisosomi, il contenuto delle vescicole dei perossisomi non è omogeneo, ma presenta aree
elettrondense (scure) centrali che risaltano sul fondo più chiaro delle vescicole.
4
CITOLOGIA 10 - Reticolo endoplasmatico, lisosomi, perossisomi, inclusioni cellulari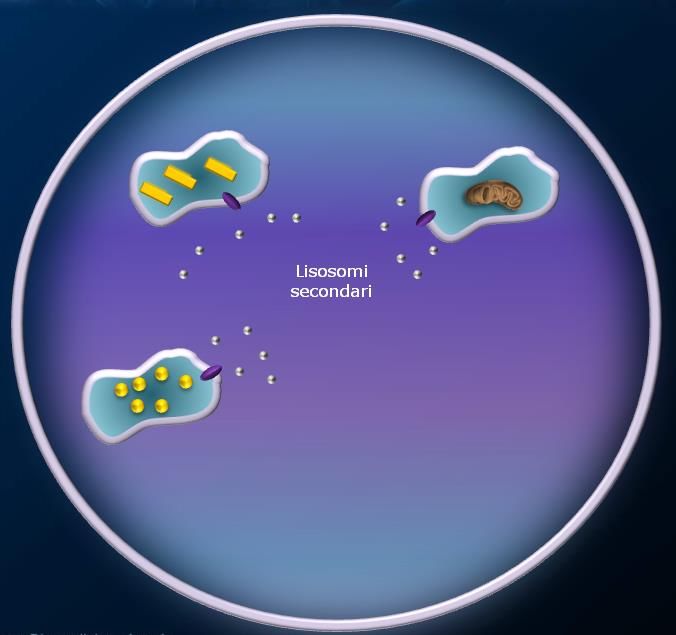
Inclusioni cellulari
Le inclusioni cellulari sono sostanze che si accumulano nel citoplasma. Esistono due tipi di inclusioni
cellulari, le inclusioni cellulari esogene e quelle endogene.
Inclusioni cellulari esogene
Sono sostanze incluse provenienti dall’esterno. Alcuni esempi sono:
carotene, una sostanza lipofila contenuta nelle carote. Se assunto in larghe quantità, può provocare
una colorazione arancione della pelle
metalli pesanti, come il piombo o l’argento
altre sostanze come l’amianto e la silice (che possono rompere le membrane lisosomali)
Inclusioni cellulari endogene
Sono sostanze prodotte dalla cellula ed immagazzinate nel citoplasma. Queste comprendono:
gocce lipidiche: si tratta di gocce di grassi che si accumulano nel citoplasma di alcune cellule (come
adipociti, cellule epatiche,…). In TEM, appaiono come gocce fortemente elettrodense, mentre in LM
sono chiare.
glicogeno: è la molecola di riserva energetica utilizzata da molte cellule del nostro organismo
(cellule epatiche, muscolari, surrenali, …). In TEM, appaiono come “rosette” elettrodense mentre in
LM sono visibili solo con particolari colorazioni.
precursori: sono sostanze momentaneamente parcheggiate nel citoplasma e che devono essere
espulse all’esterno della cellula, per svolgere una specifica funzione. Il gruppo dei precursori
comprende:
precursori di enzimi (zimogeni): enzimi in forma inattiva che, liberati fuori dalla cellula, si
attivano svolgendo la propria funzione. Alcuni esempi sono:
Tripsinogeno, un enzima proteolitico che si trova nelle cellule del pancreas.
Pepsinogeno, un enzima proteolitico che si trova nelle cellule dello stomaco.
Precursori di amilasi, enzimi glucolitici che si trovano nelle cellule del pancreas.
Altri precursori: sono molecole non enzimatiche che, una volta all’esterno della cellula, si
trasformano in altre sostanze. Esempi:
mucinogeno, abbondante nelle cellule caliciformi degli epiteli gastrointestinali e
respiratori. Esso forma la mucina, una sostanza che, a contatto con l’acqua, diventa
muco e svolge una funzione protettiva.
pigmenti: sostanze che assumono un particolare colore alla microscopia ottica. I più importanti
sono:
melanina, un pigmento marrone responsabile del colore della pelle e dei capelli.
lipofuscina, una sostanza giallognola che forma i corpi residui dei lisosomi terziari.
cristalloidi: molecole che assumono la forma di cristalli. I cristalloidi si trovano negli eosinofili
(particolari tipi di globuli bianchi) e nelle cellule di Leydig del testicolo.
5
CITOLOGIA 10 - Reticolo endoplasmatico, lisosomi, perossisomi, inclusioni cellulari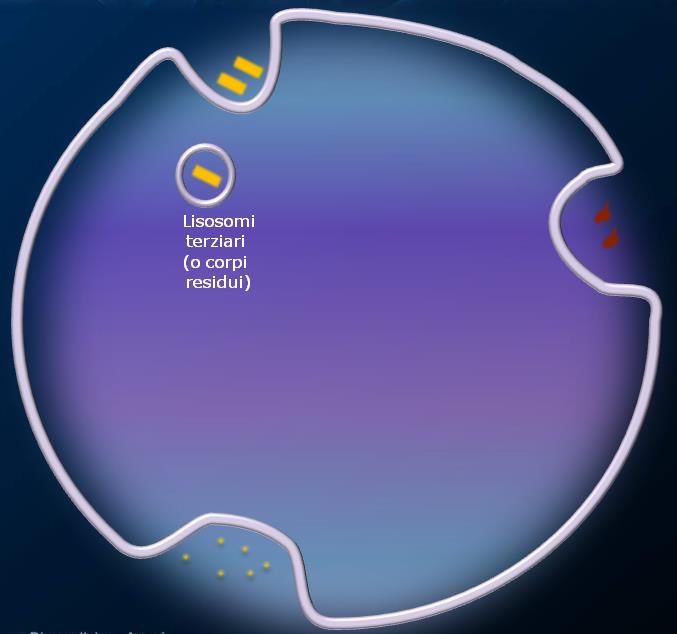
Puoi anche leggere

















































