UNIVERSITA' POLITECNICA DELLE MARCHE - UNIVPM
←
→
Trascrizione del contenuto della pagina
Se il tuo browser non visualizza correttamente la pagina, ti preghiamo di leggere il contenuto della pagina quaggiù
UNIVERSITA’
POLITECNICA
DELLE MARCHE
FACOLTA’ DI MEDICINA E
CHIRURGIA
Corso di Laurea in Medicina e Chirurgia
SINDROME METABOLICA ED EPATOPATIA
NEI PAZIENTI CON CRANIOFARINGIOMA:
epidemiologia e determinanti.
Relatore Chiar.mo Tesi di Laurea di:
Prof. Giorgio Arnaldi Chiara D’Isabella
A.A. 2020\2021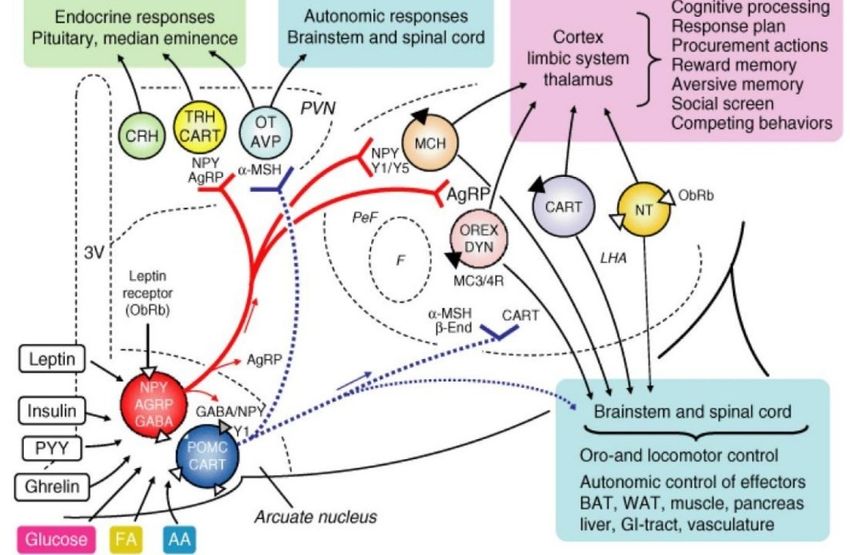
1
A nonno:
non saresti potuto venire con le gambe,
sei venuto con le ali.
2
Sommario
Introduzione: ..................................................................................................................................... 5
Ipotalamo e ipofisi ......................................................................................................................... 5
Anatomia, origine embriologica e funzione dell’ipotalamo .................................................... 5
Anatomia, origine embriologica e funzione dell’ipofisi ......................................................... 14
Omeostasi metabolica: ............................................................................................................ 19
Il craniofaringioma .......................................................................................................................... 21
Definizione ................................................................................................................................... 21
Storia ............................................................................................................................................ 22
Epidemiologia .............................................................................................................................. 23
Localizzazione .............................................................................................................................. 24
Anatomia Patologica ................................................................................................................... 24
Patogenesi ................................................................................................................................... 27
Teorie patogenetiche .............................................................................................................. 27
Mutazioni ................................................................................................................................. 28
Manifestazioni cliniche:............................................................................................................... 30
Diagnosi: ...................................................................................................................................... 31
Imaging: ....................................................................................................................................... 32
Trattamento:................................................................................................................................ 33
Comorbidita’ e sequele: .............................................................................................................. 35
Sindrome metabolica: ..................................................................................................................... 39
Definizione: .................................................................................................................................. 39
Epidemiologia: ............................................................................................................................. 39
Patogenesi: .................................................................................................................................. 39
Management: .............................................................................................................................. 41
Condizioni associate alla sindrome metabolica – la NAFLD:...................................................... 42
CONTRIBUTO SPERIMENTALE ......................................................................................................... 43
Premesse:......................................................................................................................................... 43
Obiettivi: .......................................................................................................................................... 44
Materiali e metodi: .......................................................................................................................... 44
Disegno dello studio: ................................................................................................................... 44
Pazienti e metodi: ........................................................................................................................ 45
Reclutamento dei pazienti....................................................................................................... 45
Metodi:..................................................................................................................................... 45
Analisi Statistica: ......................................................................................................................... 45
Risultati: ........................................................................................................................................... 46
3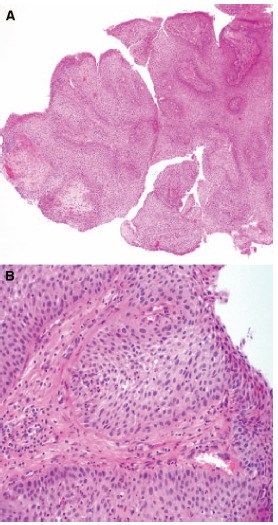
Caratteristiche dei pazienti: ........................................................................................................ 46
Genere, età e durata follow up: .............................................................................................. 46
Caratteristiche del craniofaringioma e compenso della funzione ipofisaria:............................ 46
Esordio: .................................................................................................................................... 46
Sede: ......................................................................................................................................... 47
Idrocefalo: ................................................................................................................................ 47
Danno ipotalamico ed invasione del terzo ventricolo: ........................................................... 47
Danno visivo: ........................................................................................................................... 48
Terapia antiepilettica: ............................................................................................................. 48
Terapia psicotropa:.................................................................................................................. 49
Stato di BRAF: .......................................................................................................................... 49
Neurochirurgia:........................................................................................................................ 49
Radioterapia: ........................................................................................................................... 49
Recidiva: ................................................................................................................................... 50
Ipopituitarismo: ....................................................................................................................... 51
Stratificazione dei pazienti secondo il burden di malattia:.................................................... 51
Caratteristiche della sindrome metabolica: ............................................................................... 54
Diabete mellito: ....................................................................................................................... 54
Dislipidemia: ............................................................................................................................ 54
Ipertensione arteriosa: ............................................................................................................ 55
Obesità viscerale:..................................................................................................................... 55
Componenti aggiuntivi o aggravanti: ..................................................................................... 56
Sindrome metabolica: ............................................................................................................. 56
Eventi cardiovascolari: ............................................................................................................ 57
Studio dell’epatopatia: ................................................................................................................ 57
Discussione: ..................................................................................................................................... 58
Conclusioni:...................................................................................................................................... 62
Ringraziamenti: ................................................................................................................................ 63
Bibliografia: ...................................................................................................................................... 66
4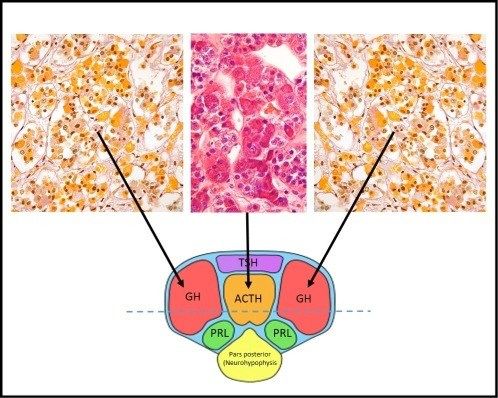
Introduzione:
Ipotalamo e ipofisi
Anatomia, origine embriologica e funzione dell’ipotalamo
L’ipotalamo è una regione della porzione ventrale del sistema nervoso centrale la cui funzione è
quella di coordinare il sistema endocrino tramite l’elaborazione di segnali provenienti da diverse
regioni del cervello, e il conseguente rilascio di ormoni a funzione regolatoria e inibitoria,
coinvolgendo nella sua azione anche l’ipofisi, la tiroide, i surreni e l’apparato riproduttivo.
È una struttura che controlla il sistema endocrino tramite proiezioni neurosecretorie magnocellulari
e parvocellulari, le prime dirette all’ipofisi posteriore mentre le seconde alla eminenza mediana.
(Shahid Z., 2022)
A livello dell’ipofisi posteriore vengono prodotte la vasopressina e l’ossitocina, implicate
nell’omeostasi osmotica e nelle funzioni riproduttive. A livello dell’ipofisi anteriore, invece,
l’ipotalamo controlla la tiroide tramite l’ormone tireotropo (TSH), la corticale del surrene tramite
l’ormone adrenocorticotropo (ACTH), i processi di crescita e omeostasi metabolica tramite
l’ormone somatotropo (GH), le gonadi tramite l’ormone luteinizzante (LH), l’ormone follicolo
stimolante (FSH) e la prolattina, coinvolta anche nella regolazione della ghiandola mammaria.
(Standring, 2009)
Inoltre, l’ipotalamo regola anche il sistema nervoso simpatico e parasimpatico, nello specifico
tramite la stimolazione della sua porzione posteriore nel primo caso e tramite la stimolazione di
quella anteriore nel secondo.
L’ipotalamo è coinvolto anche nella regolazione della pressione sanguigna, della frequenza
cardiaca, nella termoregolazione e nella regolazione dell’assunzione del cibo.
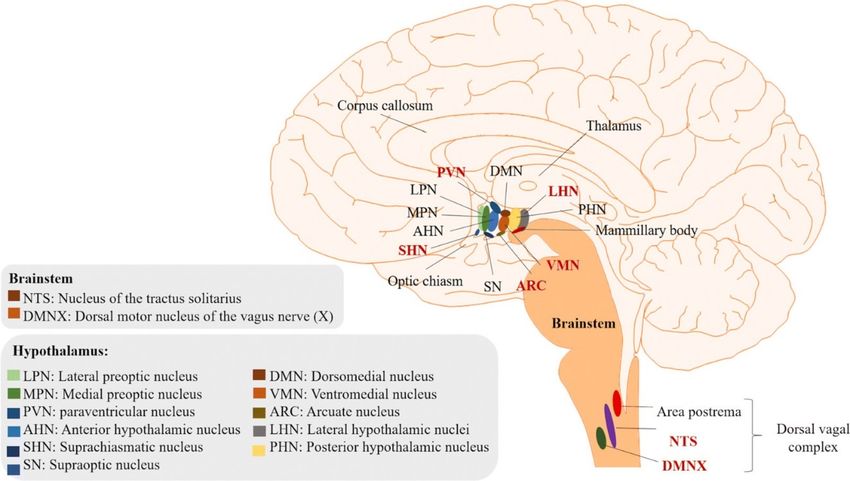
Anatomicamente, l’ipotalamo si estende dalla lamina terminale fino ai corpi mammillari, e dal solco
ipotalamico alla base dell’encefalo. Si trova sotto il talamo, al di sotto della commissura anteriore,
e davanti al subtalamo e al tegmento del mesencefalo. Lateralmente a questo, si trovano la capsula
interna ed il tratto ottico. Inferiormente all’ipotalamo si colloca l’ipofisi. Nell’adulto, questa
ghiandola non pesa più di 2-5 grammi (Standring, 2009) (Daniel, 1976)
L’ipotalamo può essere suddiviso in due regioni longitudinali, ovvero una zona mediale che
contiene la maggior parte dei nuclei ipotalamici, separata dalla zona laterale da un piano che passa
per le colonne anteriori del fornice.
5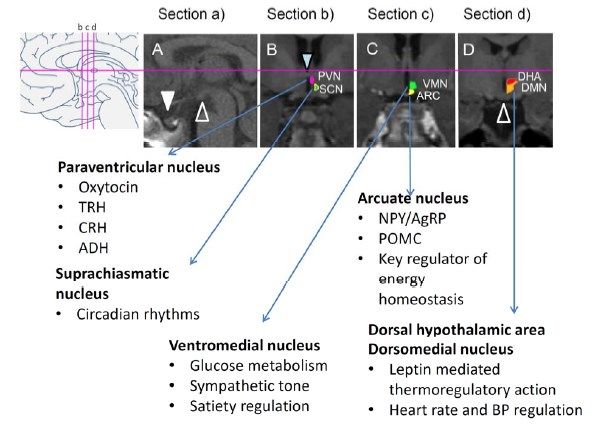
L’ipotalamo è inoltre anche suddiviso nel suo asse antero-posteriore:
• La regione preottica che si estende rostralmente al chiasma ottico e dorsalmente alla
commissura anteriore;
• La regione sovrachiasmatica che si trova al di sopra del chiasma ottico;
• La regione tuberale che si estende al di sopra ed include il tuber cinereum;
• La regione mammillare che include i corpi mammillari e i nuclei ipotalamici posteriori.
Una definizione accettata pone il limite tra la regione tuberale e quella mammillare sul piano che si
estende lungo il bordo caudale delle colonne del fornice. (Freeman, 2003)
Da un punto di vista embriologico, l’ipotalamo, il peduncolo ipofisario e l’ipofisi posteriore derivano
da un’estroflessione del cervello durante il suo sviluppo, andando a formare un’unica unità morfo-
funzionale. (Freeman, 2003)
È stato dimostrato che l’ipotalamo derivi dalla porzione più anteriore del nevrasse, sviluppandosi
dagli aspetti ventrali di due o tre segmenti rostrali del tubo neurale, le cui porzioni dorsali
andrebbero, invece, a formare il telencefalo. Il modello “prosometrico” attuale specula che una
porzione alare “pretalamica” del prosencefalo secondario vada a comprendere le porzioni
preottica, sopraottica-paraventricolare, sovrachiasmatica, anteriore, perifornicale e dorsale
dell’ipotalamo, mentre le regioni retrochiasmatica, ventrale, infundibolare e mammillare e
l’eminenza mediana derivino dalle porzioni basali degli stessi prosomeri1.
Attualmente, si sta cercando di mettere in luce i meccanismi genetici alla base dello sviluppo
ipotalamico. La proteina Sonic Hedgehod (SHH) è un morfogeno concentrazione-dipendente
prodotto dal mesoderma nevrassiale e gioca un ruolo cruciale nella sua distribuzione dorso
ventrale, regolando l’espressione di un gran numero di proteine omeobox. Questa proteina svolge
un ruolo fondamentale nella ventralizzazione durante lo sviluppo del prosencefalo.
All’interno dell’asse ipotalamo-ipofisario, la differenziazione di specifici gruppi cellulari procede in
maniera ordinata nel tempo e nello spazio, controllata da una gerarchia di fattori di trasduzione.
Nello specifico, lo sviluppo dei fenotipi cellulari presenti a livello del nucleo sovraottico,
paraventricolare, periventricolare e arcuato è regolato da Sim 1, Arnt2, Otp, Brn2 e Gsh1, ai quali si
sono aggiunti recentemente Fhx5 per i corpi mammillari, Pax6 per il tratto mammillo-talamico,
1
Prosomero: neuromero organizzato in direzione rostro-caudale di pertinenza del prosencefalo
[Nieuwenhuys R. Il sistema nervoso centrale. Springer. 2010]
6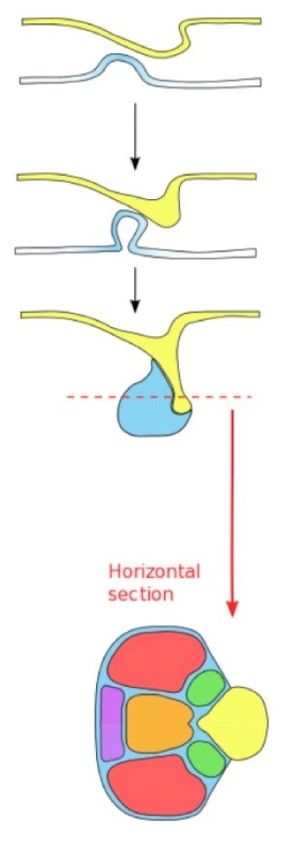
Wnt8 espresso nella regione mammillare degli embrioni umani, e SF-1, indispensabile per lo
sviluppo dei nuclei ventromediali. (Freeman, 2003)
All’interno dell’ipotalamo sono presenti regioni costituite da gruppi neuronali definite “nuclei”. Da
alcuni di questi si sviluppano connessioni ben definite dirette verso l’ipofisi posteriore. (Jenkins,
1972)
Questi nuclei sono stati tradizionalmente definiti in base ad una combinazione di criteri
embriologici, anatomici e citoarchitetturali, e permettono, ognuno con le sue particolari funzioni,
all’ipotalamo di fungere da struttura di integrazione sensitiva ed area di output motorio in grado di
mantenere l’omeostasi controllando il comportamento endocrino, autonomico e somatico.
(Freeman, 2003) (Bear MH, 2021)
[Güemes, A., Georgiou, P. Review of the role of the nervous system in glucose homoeostasis and future perspectives towards the management
of diabetes. Bioelectron Med 4, 9 (2018).]
• Nucleo paraventricolare e sovraottico: entrambi questi nuclei sono costituiti da
neuroni magnocellulare, parvocellulari e neuroni a lunga proiezione. I
magnocellulari, nello specifico, producono ossitocina e ADH, molecole che fungono
sia da ormoni che da neurotrasmettitori. Questi vengono rilasciate in maniera diretta
tramite fibre assonali a livello dei capillari della porzione posteriore dell’ipofisi.
L’ossitocina ha il compito di regolare la contrazione uterina durante il parto e la
produzione di latte durante l’allattamento, mentre l’ADH, media la traslocazione
delle acquaporine sulla superficie apicale delle cellule del tubulo contorto distale,
aumentando il riassorbimento di acqua. L’ADH, inoltre, causa vasocostrizione e
7aumento della pressione sanguigna durante l’ipovolemia, e il rilascio di fattore di von
Willebrand dalle cellule endoteliali in caso di sanguinamento. Inoltre, un sottotipo di
neuroni del nucleo paraventricolare (definiti “neuroni Nos1”, in quanto esprimono
l’ossido nitrico sintetasi 1) partecipano alla regolazione del comportamento
alimentare, proiettando al romboencefalo e a regioni del midollo implicate nel
controllo dell’assunzione del cibo e della spesa energetica. Inoltre, sia l’ADH che
l’ossitocina, agendo da neurotrasmettitori centrali, sono coinvolti nella formazione
del legame di coppia. (Freeman, 2003) (Bear MH, 2021) (Cheng Qin, 2018) (Sutton,
2014)
• “L’area preottica”: il nucleo preottico presente in quest’area coadiuva il nucleo
anteriore nella regolazione la temperatura corporea promuovendo lo scambio di
calore con l’esterno, andando a diminuire il tono simpatico a livello dei muscoli
scheletrici, aumentandolo invece a livello della pelle, provocando la dilatazione dei
capillari.
L’area preottica in generale sembra inoltre svolgere la sua azione andando anche a
regolare la termogenesi a livello del tessuto adiposo bruno. Questa associazione ha
portato allo sviluppo di studi ulteriori, i quali hanno speculato, e successivamente
confermato, un suo compito anche nell’omeostasi del peso corporeo e sulla spesa
energetica. Anche la regolazione del ritmo sonno-veglia promossa dall’OVLT sembra
essere connessa all’omeostasi del peso corporeo, tramite connessioni al sistema
oressina/ipocretina situato nell’area ipotalamica laterale che, oltre ad essere
implicato nello stimolare e mantenere il sonno, promuove anche l’attività motoria e
il rewarding legato al cibo. A questo si aggiungono anche connessioni tra l’area
preottica con l’organo subfornicale e l’OVLT, implicate nell’omeostasi osmotica: la
regolazione della sete è infatti strettamente connessa a quella della fame ed una
disregolazione a livello della prima porta a modificazioni della seconda e viceversa.
Infine, l’area preottica comunica con il nucleo premammillare ventrale per la
regolazione dell’attività sessuale. Anche quest’ultima sembra essere influenzata dal
peso, tramite neuroni che presentano sia recettori per la leptina che l’enzima ossido
nitrico sintetasi presenti a livello di entrambi i nuclei. Il cross-talk tra questi nuclei va
quindi a regolare l’espressione dei releasing hormones delle gonadotropine e, in
ultima istanza, l’attività sessuale. Inoltre, individui di sesso maschile e femminile
presentano una differente distribuzione degli estrogeni a livello del nucleo preottico,
influenzando il comportamento sessuale e materno. (Bear MH, 2021) (Freeman,
2003) (Yu S, 2018) (Rebecca L. Leshan, 2014)
8• Nucleo sovrachiasmatico: il nucleo sovrachiasmatico è il pacemaker circadiano
primario nei mammiferi, regolando la secrezione ormonale e il comportamento
diurno tramite l’input luminoso proveniente dalla retina. L’attività dei fattori di
trascrizione che oscilla nelle 24 ore assieme la modificazione della frequenza dei
potenziali d’azione incrementano l’attività motoria durante il giorno e la
diminuiscono durante la notte (ad esempio, il cortisolo ha il suo picco al sorgere del
sole, mentre l’ormone della crescita a mezzanotte. Molti degli infarti accadono la
mattina presto a causa del picco del cortisolo come ormone dello stress che, tra le
altre cose, aumenta la pressione sanguigna). Questo nucleo è composto da due parti,
una regione core e un guscio. Il core presenta al suo interno cellule che esprimono
VIP (ormone vasoattivo intestinale) e GRP (peptide di rilascio della gastrina), mentre
nel guscio sono presenti neuroni che esprimono AVP (arginina-vasopressina). Nella
regione core il VIP aumenta nei periodi di buio, mentre il GRP durante i periodi di
luce ed è attivato in maniera specifica da impulsi diretto provenienti dalle cellule
gangliari fotosensibili della retina tramite il tratto retino-ipotalamico (RHT). I neuroni
che esprimono AVP a livello della guscio, invece, proiettano al nucleo
paraventricolare, coordinando il ritmo circadiano della nutrizione. Inoltre, proiettano
anche ai neuroni implicati nel controllo della sete presenti a livello dell’OVLT. (Bear
MH, 2021) (Freeman, 2003) (Morrison, 2022)
• Nucleo ventromediale, dorsomediale e area laterale dell’ipotalamo:
Il nucleo ventromediale è direttamente connesso al comportamento alimentare. È in
grado di percepire il senso di sazietà, portando a una diminuzione dell’alimentazione.
La distruzione di quest’area porta ad iperfagia, come quella che è possibile vedere
nella sindrome di Prader-Willi. Il meccanismo attraverso il quale questo nucleo è in
grado di regolare il peso corporeo, l’assunzione degli alimenti, l’energia e l’omeostasi
del glucosio è collegata alla presenza dei LEPRs (recettori della leptina e i suoi affini);
questi recettori sono espressi anche dal nucleo arcuato a livello dei neuroni POMC e
AgrP. (Choi, 2013) Alcuni studi hanno messo in luce che la stimolazione del nucleo
ventromediale aumenta l’utilizzo del glucosio sia tramite la glicogenolisi epatica che
aumentando l’uptake di glucosio da parte dei tessuti periferici, come il tessuto
adiposo bruno e il muscolo sia scheletrico che cardiaco. Lesioni di questo nucleo
portano a un aumento dei livelli di insulina e una diminuzione di quelli di glucagone.
(Khodai, 2021)
L’area laterale dell’ipotalamo, al contrario, percepisce il senso di fame e induce lo
stimolo ad alimentarsi. La distruzione di quest’area porta all’anoressia. Inoltre, è
9interconnessa con tutti i maggiori sistemi diretti in uscita dal sistema nervoso
centrale, tra cui il sistema nervoso autonomo tramite il tronco encefalico, il midollo
spinale e il nucleo paraventricolare, l’asse neuroendocrino tramite il nucleo
paraventricolare ed altri nuclei ipotalamici, le aree di elaborazione cognitiva, la
memoria e le aree di valenza emozionale tramite la corteccia, il talamo e il sistema
limbico; per tale motivo, questa area è adatta ad orchestrare stimoli ambientali
(vista, odorato, gusto, temperatura, cognizione) e metabolici (glucosio nel sangue,
ormoni circolanti, infiammazione) in una risposta appropriata da parte del sistema
autonomico, degli assi neuroendocrini e del comportamento. (Qualls-Creekmore E,
2018)
Il nucleo dorsomediale, invece, controlla la rabbia, la quale può essere scatenata, in
relazione alla vicinanza con gli altri due nuclei, della mancata sensazione di sazietà.
Questo nucleo è inoltre implicato nelle risposte autonomiche allo stress,
nell’alimentazione e nel rilascio circadiano di corticosteroidi. Ulteriormente
possiamo dire che, presenta delle proiezioni efferenti dirette al nucleo
ventromediale e a due distinte porzioni del nucleo paraventricolare, ovvero quelle
costituite dai neuroni parvocellulari e magnocellulari.
(Bear MH, 2021) (Freeman, 2003) (Chou TC, 2003) (Kalra SP, 1999)
• Nucleo arcuato: è uno dei più importanti nuclei che rilasciano ormoni tramite terminali
assonali diretti al sistema venoso portale ipotalamo-ipofisario, che controlla il rilascio
ormonale da parte dell’ipofisi anteriore. Dal punto di vista metabolico, il nucleo
arcuato presenta neuroni che producono peptidi oressizzanti (NPY), oppioidi
(dinorfine), peptidi derivati da POMC (beta-endorfine e galanina), e gli amminoacidi
(glutammato e GABA). I peptidi anoressizzanti, alfa-MSH e AgrP, sono strettamente
correlati a quelli oressizzanti appena citati: alfa-MSH deriva anche questo da POMC ed
è prodotto insieme alla beta-endorfina, mentre l’mRNA di AgrP è coespresso assieme
a quello di NPY.
I peptidi CART (trascritto regolato da cocaina e anfetamina) sono stati riconosciuti
come importanti segnali anoressizzanti.
Il nucleo arcuato non possiede la barriera ematoencefalica e si trova quindi in una
posizione strategica per avvertire segnali periferici, come steroidi gonadici e surrenali,
oltre che grandi peptidi, come leptina e insulina, ed altri segnali trasportati dal liquido
cefalorachidiano all’interno del sistema cerebro-vascolare. (Kalra SP, 1999)
Tutte queste molecole sono espresse da popolazioni neuronali discrete che mediano
sia l’assunzione di cibo che la spesa energetica. Queste popolazioni sono due e sono i
10neuroni oressizzanti NPY/AgrP (neuropeptide Y/ agouti-related peptide) e i neuroni
anoressizzanti POMC/CART (pro-opiomelanocortin/cocaine-amphetamine related
transcript). L’attivazione dei primi promuove i processi anabolici che portano ad un
aumento dell’assunzione di cibo, mentre una loro lesione porta a perdita di peso. Al
contrario, l’attivazione dei secondi favorisce i processi catabolici, riducendo l’appetito
e l’assunzione di cibo. Recentemente, una nuova popolazione di neuroni è stata
scoperta a livello del nucleo arcuato, i neuroni TH (tirosina idrossilasi) che hanno
effetto oressizzante. (Güemes, 2018)
• Connessioni ipotalamo-ipofisi: molti nuclei dell’ipotalamo concorrono alla
regolazione del rilascio di ormoni da parte dell’ipofisi; questi producono releasing-
hormones che sono trasportati a livello ipofisario e stimolano specifiche cellule a
rilasciare il loro secreto.
Il CRH (corticotropin-releasing hormone) provoca il rilascio di ACTH (ormone
adrenocorticotropo) dai pituiciti ai capillari; questo ormone viaggia attraverso la
circolazione sistemica e stimola la corteccia surrenalica (Zona reticularis) a produrre
cortisolo.
Il TRH (thyrotropin-releasing hormone), invece, porta alla secrezione di TSH il quale
aumenta la sintesi e il rilascio degli ormoni tiroidei che sono in grado di regolare il
metabolismo. L’ipotalamo, infatti, è in grado di percepire la presenza delle riserve
energetiche del corpo tramite recettori per la leptina e, nel caso in cui queste
dovessero essere basse, è in grado di diminuire il metabolismo, diminuendo il rilascio
di ormoni tiroidei. Il rilascio pulsatile di GnRH (gonadotropin-releasing hormone)
aumenta i livelli di LH (ormone luteinizzante) e FSH (ormone follicolostimolante); l’LH
stimola i testicoli maschili a produrre testosterone e le ovaie femminili a produrre
estrogeno, ormoni che portano allo sviluppo dei caratteri sessuali secondari sia negli
individui di sesso maschile che femminile. Un picco di LH causa l’ovulazione. L’FSH,
invece, stimola la spermatogenesi maschile e la maturazione oocitaria femminile.
Il GHRH (growth hormone releasing hormone) porta al rilascio di GH (ormone della
crescita), che stimola il metabolismo e la crescita tissutale. (Bear MH, 2021) (Freeman,
2003)
I neuroni che producono TRH, che stimola il rilascio di TSH e prolattina sono localizzati
soprattutto a livello dell’ipotalamo laterale; i neuroni che rilasciano dopamina, che
inibisce il rilascio di prolattina da parte dell’ipofisi si trovano soprattutto a livello della
regione mediale del nucleo arcuato; i neuroni che invece producono CRH, stimolano la
produzione ipofisaria di ACTH ed endorfine, sono posti nella porzione laterale del
11nucleo paraventricolare; quelli che producono GHRG si trovano nella porzione laterale
del nucleo arcuato e sono inibiti da neuroni che producono GHIH, situati nel nucleo
paraventricolare; infine i neuroni che producono GNRH sono localizzati nell’area
preottica mediale. (Giuseppe Musumeci, 2015)
• Corpi mammillari: sono inseriti all’interno del sistema limbico come parte del circuito
di Papez e sono implicati nella formazione dei ricordi. Una lesione bilaterale dei corpi
mammillari porta alla sindrome di Wernicke Korsakoff, caratterizzata da amnesia
anterograda e talvolta anche retrograda. (Bear MH, 2021) (Freeman, 2003)
Regione Zona Nucleo Caratteristiche
funzionali
Preottica Mediale OVLT (organum Osmorecezione e
vasculosum della chemorecezione,
lamina terminale) ritmo sonno-veglia
Organo subfornicale Sensore
dell’angiotensina
circolante, centro
della sete
N. preottico Dissipazione di calore
N. intermedio Nucleo dismorfico
sessuale, regolazione
della funzione
riproduttiva
Sopraottica Mediale N. anteriore Dissipazione del
calore, eccitazione del
parasimpatico
N. sovrachiasmatico Orologio biologico,
ritmo circadiano
N. paraventricolare Secrezione ADH e
ossitocina
Entrambe N. sovraottico Osmorecezione,
secrezione ADH e
ossitocina
12Tuberale Mediale N. ventromediale Centro della sazietà;
causa aggressività se
distrutto
N. dorsomediale Non definite,
controllo della rabbia
Zona periventricolare Controllo dell’ipofisi
anteriore
N. infundibolare Controllo dell’ipofisi
(arcuato) anteriore, regolazione
di peso e appetito
Entrambe N. tuberomammillare Risveglio e veglia
Laterale N. tuberali laterali Non definita
Area laterale Centro della fame
ipotalamica
Mammillare Mediale Corpi mammillari Memoria a breve
termine, esperienza
emozionale
N. posteriore Conservazione del
calore; eccitazione del
simpatico
/ N. premammillare Regolazione della
ventrale funzione riproduttiva
Tutte le zone Laterale Area ipotalamica Nuclei interstiziali del
laterale prosencefalo
(Bear MH, 2021) (Yu S, 2018)
L’ipotalamo riceve come input da un network ampiamente distribuito di fibre che giungono dal
prosencefalo, dal troncoencefalico e dal midollo spinale. Molte di queste connessioni sono
reciproche e giungono a molteplici destinazioni intra-ipotalamiche.
Il fascio prosencefalico mediale attraversa l’ipotalamo laterale, si continua rostralmente con le aree
settali e caudalmente con il tegmento del tronco encefalico, e presenta nuclei interstiziali al suo
interno. Il fascicolo longitudinale dorsale connette i nuclei della zona periventricolare mediale con
il grigio periacqueduttale. L’amigdala, invece, invia due principali proiezioni all’ipotalamo, ovvero la
stria terminale e il tratto amigdalo-fugale ventrale.
13Gli input provenienti dalla formazione ippocampale arrivano primariamente tramite il fornice,
fornendo connessioni ai nuclei della zona mediale prima di terminare nei corpi mammillari.
Altri importanti input sono quelli della retina, deli organi circumventricolari, della corteccia basale
frontale e dell’insula. (Freeman, 2003)
Anatomia, origine embriologica e funzione dell’ipofisi
L’ipofisi è un piccolo organo di forma ovale con un diametro trasverso di circa 12 mm e un diametro
anteroposteriore di 8 mm, ed un peso di circa 500 mg. Il lobo anteriore dell’ipofisi è normalmente
più piccolo negli uomini rispetto alle donne, e nelle nullipare piuttosto che nelle multipare.
È localizzato nella sella turcica, una struttura concava che si trova a livello dell’aspetto superiore
dell’osso sfenoide.
Superiormente è
coperta dal diaphragma
sellae ed è in rapporto
con l’ipotalamo ed il
chiasma ottico, mentre
lateralmente si possono
trovare i seni cavernosi
che contengono le
arterie carotidee interne
(Asa S L. Tumors of the Pituitary Gland. AFIP Atlas of Tumor e i nervi oculomotore,
Pathology, 2011) trocleare, abducente e
la prima divisione dei nervi trigeminali. Inferiormente, invece, è in contatto con la parte posteriore
del seno sfenoidale. (Asa S L. Tumors of the Pituitary Gland. AFIP Atlas of Tumor Pathology, 2011)
(Larkin S)
L’ipofisi presenta due porzioni, ovvero l’adenoipofisi (ipofisi anteriore) e la neuroipofisi (ipofisi
posteriore).
✓ L’adenoipofisi costituisce circa l’80% dell’intera ghiandola pituitaria; è composta da tre
parti: la pars distalis, la pars intermedia e la pars tuberalis. La pars distalis è la porzione più
ampia della ghiandola ed è anche conosciuta con il nome di pars glandularis. La pars
intermedia è rudimentale nell’ipofisi umana, è la rimanenza della porzione posteriore della
tasca di Rathke. La pars tuberalis si sviluppa verso l’alto e circonda la parte inferiore del
peduncolo ipofisario, ed è anche conosciuta come pars infundibularis. (Asa S L. Tumors of
the Pituitary Gland. AFIP Atlas of Tumor Pathology, 2011)
14Istologicamente, l’adenoipofisi è caratterizzata da acini ben demarcati che contengono
diverse cellule secernenti ormoni, ben visibile attraverso preparazioni a base di reticolina.
Le cellule corticotrope
sono fortemente basofile
(PAS positive), le
somatotrope e lattotrope
sono acidofile, mentre le
gonadotrope e le
tireotrope sono spesso
basofile o cromofobe. In
un adulto normale circa il
10% delle cellule
dell’ipofisi sono basofile, il
40% acidofile e il 50%
cromofobe. (Larkin S)
È stato messo in luce che le zone laterali della ghiandola contengano soprattutto cellule
somatotrope e lattotrope, la zona centrale invece soprattutto corticotrope, mentre la
porzione anteriore presenta soprattutto cellule tireotrope; le gonadotrope sono per lo più
diffuse. (Larkin S)
✓ La neuroipofisi è composta da fibre nervose provenienti da nuclei ipotalamici che
proiettano verso il basso; dà origine all’eminenza mediana (o infundibolo), al peduncolo
ipofisario e al lobo posteriore. (Asa S L. Tumors of the Pituitary Gland. AFIP Atlas of Tumor
Pathology, 2011)
L’ipofisi è circondata dalla dura madre. Il diaphragma sellae è un riflesso della dura madre che va a
costituire il tetto della sella turcica presenta un piccolo buco per il peduncolo ipofisario che
connette ipofisi a ipotalamo, la cui funzione è quella di proteggere l’ipofisi dalla pressione del
liquido cerebro-spinale.
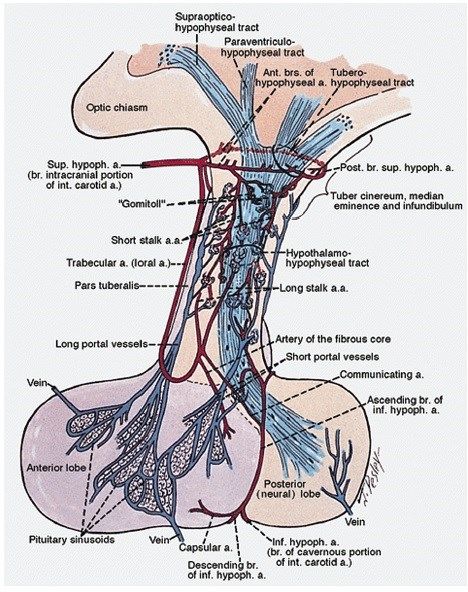
La vascolarizzazione ipofisaria comprende un complesso sistema portale che origina dall’ipotalamo
e che trasporta al suo interno ormoni stimolatori ed inibitori fino alle cellule dell’adenoipofisi,
giocando quindi un ruolo importante nella sua secrezione ormonale. La vascolarizzazione arteriosa
dell’eminenza mediana e dell’ipofisi posteriore deriva da due o tre arterie che derivano a loro volta
dalla porzione intracranica delle arterie carotidi interne, ovvero le arterie ipofisarie superiore,
media e inferiore. L’arteria ipofisaria superiore crea due plessi, uno esterno ed uno interno:
l’esterno è composto da piccole arterie che circondano la porzione superiore del peduncolo e che
15danno origine ad un groviglio di
capillari; l’interno, invece, forma i
“gomitoli”, strutture vascolari
uniche di 1-2 mm di lunghezza e 0,1
mm di spessore, composte da una
arteria muscolare centrale
circondata da spirali di capillari; le
arteriole forniscono sangue ai
capillari attraverso piccoli orifizi
circondati da sfinteri muscolari. Il
flusso attraverso queste complesse
strutture procede poi, attraverso il
sistema portale, ai capillari
dell’adenoipofisi. Nonostante la loro
funzione non sia stata ancora ben
definita, la complessità di questi
gomitoli suggerisce che possano
essere in grado di regolare il flusso
(Asa S L. Tumors of the Pituitary Gland. AFIP Atlas
of Tumor Pathology, 2011) sanguigno all’ipofisi anteriore,
andando ad influenzare il trasporto degli ormoni ipotalamici regolatori.
Le arterie ipofisarie inferiori penetrano la sella turcica appena al di sotto del suo diaframma e
forniscono sangue alla capsula ipofisaria, al lobo posteriore e alla porzione inferiore del peduncolo
ipofisario. All’interno del solco intra-lobare si dividono in tronchi ascendenti e discendenti, andando
a formare un circolo arterioso attorno alla neuroipofisi. I capillari della neuroipofisi sono fenestrati
e si trovano al di fuori della barriera ematoencefalica.
Precedentemente si pensava che i lunghi vasi portali che originavano dall’infundibolo portassero
dal 70 al 90 % del sangue diretto all’ipofisi, mentre il 10-30% rimanente originasse da piccoli vasi
portali che connettono il processo infundibolare all’adenoipofisi; al giorno d’oggi si pensa, invece,
che il flusso sanguigno passi attraverso i capillari della neuroipofisi, risultando in una commistione
di sangue proveniente da diversi vasi portali. L’adenoipofisi riceve la maggior parte del suo flusso
sanguigno da vasi portali che derivano dalla neuroipofisi, ma presenta anche sangue arterioso
proveniente da due diramazioni della arteria ipofisaria inferiore: l’arteria capsulare, che
vascolarizza il tessuto connettivo della capsula ipofisaria, e l’arteria del nucleo fibroso.
16Il drenaggio venoso della ghiandola pituitaria è diretto verso i seni cavernosi e da qui verso i seni
petrosi inferiori; il volume del sistema venoso che si diparte dall’adenoipofisi e neuroipofisi verso i
seni cavernosi è considerevolmente minore rispetto alla vascolarizzazione in entrata.
Per quel che concerne la componente nervosa che serve la ghiandola ipofisaria, questa è
estremamente particolare e cruciale per le funzioni della ghiandola; va anche ricordato che
l’adenoipofisi non presenta alcuna afferenza nervosa tranne che poche fibre simpatiche associate
ai capillari in modo da regolare il flusso sanguigno all’adenoipofisi. D’altra parte, il lobo posteriore
è quasi unicamente composto da assoni e fibre nervose che derivano dall’ipotalamo, importanti per
la secrezione dei due ormoni caratteristici di questa zona, ovvero l’ossitocina e la vasopressina,
oltre che il trasporto dei peptidi ipotalamici che regolano la funzione dell’adenoipofisi. Il tratto
ipotalamo-ipofisario è costituito principalmente da fibre nervose derivanti dal nucleo sovraottico e
paraventricolare che trasportano vasopressina e ossitocina a livello della porzione posteriore
dell’ipofisi dove sono rilasciati, a livello dei capillari.
Il tratto tubero-infundibolare origina da neuroni neurosecretori che producono ormoni ipofisiotropi
e proietta a diversi nuclei dell’eminenza mediana dove gli ormoni sono rilasciati a livello del sistema
vascolare ipofisario (Larkin S) (Asa S L. Tumors of the Pituitary Gland. AFIP Atlas of Tumor Pathology,
2011).
ORMONE ORIGINE FUNZIONE
Ossitocina Dai neuroni magnocellulari dei Nelle donne porta alla
nuclei sovraottico e paraventricolare contrazione della parete uterina
dell’ipotalamo; e all’eiezione di latte in risposta
all’estradiolo, allo stimolo della
sanzione; mentre, negli uomini,
riduce la sintesi di steroidi
testicolari.
Vasopressina: Dai neuroni magnocellulari dei Risposta agli osmocettori
nuclei sovraottico e paraventricolare sensibili alle concentrazioni di
dell’ipotalamo; sodio extracellulare; Aumento
del riassorbimento renale di
acqua dai tubuli collettori;
Riduzione della frequenza
respiratoria in risposta alla
disidratazione; Aumento della
pressione sanguigna tramite la
contrazione arteriolare.
Corticotropin- Dai nuclei ipotalamici parvocellulari: Azione sulle cellule ipofisarie
releasing Hormone arcuato e paraventricolare mediale; corticotrope che rilasciano ACTH
e beta-endorfina.
Gonadotropin- Dai nuclei ipotalamici parvocellulari: Azione sulle cellule ipofisarie
releasing Hormones arcuato e paraventricolare mediale; gonadotrope che rilasciano FSH
ed LH.
17Growth hormone Dal nucleo ipotalamico
Azione sulle cellule ipotalamiche
releasing Hormone parvocellulare arcuato; GH secernenti.
Growth hormone- Dai nuclei ipotalamici parvocellulari:
Oltre che inibire le cellule GH
inhibiting hormone (o arcuato e paraventricolare mediale;secernenti, la somatostatina è
somatostatina); coinvolta in molte funzioni
cerebrali, come il
comportamento,
l’apprendimento, la secrezione
endocrina, la proliferazione
cellulare e l’apoptosi.
Dopamina Dal nucleo ipotalamico Inibizione della secrezione di
parvocellulare arcuato; prolattina nelle cellule
lattotrope.
Thyrotropin Dal nucleo ipotalamico Stimolazione del TSH;
parvocellulare ventromediale, Stimolazione della sintesi e
Releasing Hormone
dorsale e paraventricolare; rilascio di prolattina.
[
(Giuseppe Musumeci, 2015)]
Per quel che concerne l’embriologia l’adenoipofisi deriva
dalla tasca di Rathke, ovvero una invaginazione
dell’ectoderma del cavo orale primitivo (Asa S L. Tumors
of the Pituitary Gland. AFIP Atlas of Tumor Pathology,
2011), ha un’origine epiteliale, a differenza della
neuroipofisi, che invece deriva invece dall’ectoderma
neurale (Larkin S).
Durante la terza e quarta settimana di gestazione,
l’ectoderma si ispessisce e si comincia a invaginare;
mentre, durante la quinta settimana la tasca di Rathke
diventa un lungo tubo con un lume stretto e una parete
spessa composta da epitelio cuboide stratificato. Alla
sesta settimana poi, la connessione con l’orofaringe viene
totalmente obliterata e la tasca di Rathke entra in
contatto diretto con l’estensione dell’ipotalamo che dà
origine all’infundibolo; questi due tessuti vengono poi
avvolti dalla cartilagine dell’osso sfenoide che li separa
dallo stomodeo, mentre la sella turcica si forma attorno
alla settima settimana. A questo punto la base della tasca
di Rathke si separa completamente dall’epitelio orale
(Asa S L. Tumors of the Pituitary Gland. AFIP Atlas of
Tumor Pathology, 2011).
(Larkin S)
18Le cellule della porzione anteriore della tasca vanno contro a una importante proliferazione che
andrà a formare il lobo anteriore, mentre la parete posteriore subirà una proliferazione più lenta,
andando a formare il lobo intermedio (Larkin S). Il lobo anteriore, durante la sua crescita, si estende
lateralmente e segue una crescita triradiata: le zone laterali diventano le ali laterali della ghiandola
nell’adulto, mentre la porzione mediale diventa il cuneo mucoide anteromediale (Asa S L. Tumors
of the Pituitary Gland. AFIP Atlas of Tumor Pathology, 2011).
Durante la crescita e lo sviluppo della porzione anteriore della ghiandola, la pars nervosa si sviluppa
sottoforma di un’evaginazione diretta verso il basso del diencefalo ventrale (Larkin S); il lume di
questa tasca si chiude andando a formare il peduncolo ipofisario, mentre la porzione superiore
forma un recesso sul pavimento del terzo ventricolo detto eminenza mediana (Nussey S, 2001).
Il limite tra la tasca di Rathke e la pars nervosa diviene mano a mano indistinto ed è formato da
resti del lume obliterato, ovvero cavità cistiche circondate da epitelio cuboidale o colonnare.
Lo sviluppo e la citodifferenziazione sono regolati da specifici fattori di trascrizione, tra i quali Rpx,
Pax-6, Ptx-1 e Ptx-2 e sono tutti necessari nell’organogenesi precoce della ghiandola pituitaria; oltre
questi fattori di trascrizione, sono importanti anche altri fattori regolatori, tra cui ormoni trofici
ipotalamici, sistemi a feedback ormonali provenienti da organi target, e fattori di crescita (Asa S L.
Tumors of the Pituitary Gland. AFIP Atlas of Tumor Pathology, 2011).
Omeostasi metabolica:
Uno dei principali meccanismi attraverso cui l’ipotalamo è coinvolto nella omeostasi energetica è
soprattutto la regolazione dell’assunzione del cibo; tra i nuclei, presenti a livello ipotalamico, il
nucleo arcuato è uno dei principali regolatori dell’appetito, in quanto al suo interno sono presenti
i neuroni POMC (pro-opiomelanocortina) e CART (trascritto regolato da anfetamina e cocaina), che
hanno il compito di diminuire l’appetito e di aumentare la spesa energetica, oltre che i neuroni
AgrP (proteina correlata ad Agouti) e NPY (neuropeptide Y), che aumentano l’appetito e
diminuiscono la spesa energetica. Tutti questi neuroni sono modulati da segnali ormonali periferici
(tra i quali glucosio, acidi grassi ed amminoacidi (Lenard, 2008)), e a sua volta il nucleo arcuato
comunica con altri neuroni oressizzanti ed anoressizzanti presenti in altri nuclei dell’ipotalamo
(Farr, 2016). I più importanti siti verso cui proietta il nucleo arcuato sono popolazioni di neuroni di
secondo ordine a livello dell’area ipotalamica laterale o peri-fornicale, e il nucleo paraventricolare
dell’ipotalamo; input provenienti dai neuroni NPY/AgrP sono controbilanciati da input provenienti
dai neuroni POMC e questa informazione integrata, inerente lo stato metabolico, viene connessa
ad altre informazioni provenienti da altre aree del cervello (aree associate alla ricompensa, alla
motivazione, al linguaggio, alla memoria, aree associate alla percezione vagale, aree associate alla
19sensibilità viscerale, aree associate alla coordinazione sensitivo-motoria ed aree associate alla
veglia (Lenard,
2008).
I neuroni presenti
a livello dell’area
ipotalamica
laterale, che
ricevono input dal
nucleo arcuato,
presentano al loro
interno diversi
neuropeptidi
correlati all’alimentazione, tra cui oressina/ipocretina, (Lenard, 2008)
MCH (melanin concentrating hormone), CART,
neurotensina ed istamina; questi neuroni di secondo ordine proiettano in maniera ampia a tutto il
cervello dalla corteccia fino al midollo spinale.
I neuroni di secondo ordine del nucleo paraventricolare sono associati alle funzioni autonomiche e
neuroendocrine; i neuroni che rilasciano tireotropina e corticotropina ricevono input diretti da
entrambe le tipologie di neuroni del nucleo arcuato, andando, successivamente, a regolare l’asse
tireotropo e corticotropo (Lenard, 2008).
Per quel che concerne i neuroni che esprimono AgrP, questi presentano effetti oressizzanti quando
attivati da ormoni della fame come la grelina, mentre ormoni della sazietà, come PYY, GLP-1,
leptina, insulina e colecistochinina, inibiscono questi neuroni. Gli AgrP sono neuroni che si attivano
in particolar modo con il digiuno; infatti, sono connessi ad altri circuiti neuronali correlati alla
sopravvivenza.
I neuroni che esprimono POMC sono attivati dalla leptina e dalla insulina, ed inviano un segnale
anoressizzante ad alti nuclei ipotalamici e ad altre aree del cervello tramite l’alfa-MSH (Laura van
Iersel, 2019).
Una condizione particolare è rappresentata dal fatto che le popolazioni neuronali che esprimono
POMC/CART ed NPY presentino una inibizione a livello locale: i neuroni NPY producono anche GABA
e, quando attivati, vanno ad inibire i neuroni POMC sia tramite recettori GABAa che recettori NPY-
1. Questo assetto è una sorta di servo meccanismo che mantiene lo stimolo all’alimentazione
sempre attivo in assenza dell’inibizione da parte dei neuroni NPY su quelli POMC. (Farr, 2016)
20Dal nucleo paraventricolare e dall’area laterale dell’ipotalamo partono proiezioni rivolte verso il
locus coeruleus (che controlla il sistema nervoso simpatico) e al nucleo motore dorsale del vago
(che controlla il nervo vago, ovvero uno dei principali costituenti del sistema nervoso
parasimpatico).
Negli stati di buona riserva energetica, gli elevati livelli di leptina e di insulina fanno si che il braccio
anoressizzante ipotalamico attivi il sistema nervoso simpatico; la stimolazione dei recettori beta-2
adrenergici provoca l’aumento dell’espressione di numerosi geni a livello dei muscoli scheletrici,
che promuovono la biogenesi mitocondriale, la glicogenolisi, la termogenesi ed in generale
incrementano tutti i sistemi atti a incrementare la spesa energetica. L’attivazione, invece, dei
recettori alfa-2 stimola la formazione delle proteine Gi, che inibiscono l’adenilato ciclasi,
diminuiscono il cAMP e mantengono i canali del potassio in una configurazione aperta che
determina la presenza di un potenziale di membrana negativo a riposo, in maniera tale da ridurre
la secrezione pancreatica di insulina e di conseguenza l’accumulo di energia sottoforma di tessuto
adiposo. Infine, l’attivazione del simpatico stimola i recettori beta-3 adrenergici sugli adipociti
promuovendo la lipolisi.
Al contrario, nelle situazioni di digiuno, le leptina e l’insulina sono basse, ciò porta ad una riduzione
del tono del simpatico, della termogenesi a livello del muscolo scheletrico e della lipolisi a livello di
quello adiposo; inoltre, l’area laterale del talamo ed il nucleo paraventricolare mandano proiezioni
tramite il fascicolo longitudinale mediale al nucleo motore dorsale del vago. Tramite il suo effetto,
all’interno del canale alimentare il vago promuove la peristalsi e l’assorbimento del cibo, a livello
cardiaco riduce la frequenza cardiaca e il consumo di ossigeno a livello del miocardio, a livello degli
adipociti promuove l’aumentata azione della lipoprotein-lipasi, per convertire i substrati energetici
in tessuto adiposo, infine, tramite la sua azione sulle cellule beta pancreatiche, il vago aumenta
l’ipersecrezione insulinica post-prandiale in risposta al pasto (Lusting, 2011).
Il craniofaringioma
Definizione
Il craniofaringioma è un tumore epiteliale raro, benigno e a lenta crescita, localizzato soprattutto
nella regione sellare, parasellare e sovrasellare del Sistema Nervoso Centrale (Garnett MR, 2007)
(Zoicas F, 2012), derivante dai resti della cosiddetta “Tasca di Rathke”.
Questo tumore presenta un basso grade istologico (WHO I) (WHO, World Health Organization
Classification of Tumours of the Central Nervous System, 2021); in passato, era stato
istologicamente distinto in due varianti, ovvero l’adamantinomatoso e il papillare (Hamblin R,
2021). Nel 2021, invece, la WHO Classification of Tumors of the Central Nervous System ha definito
21queste due tipologie come entità tumorali distinte viste le loro differenze dal punto di vista
epidemiologico, oltre che le loro caratteristiche radiografiche, alterazioni genetiche e profili di
metilazione (Louis, 2021). Questa distinzione appare ancor più chiara se si considera la distribuzione
bimodale di questi tumori tra le fasce d’età dei soggetti affetti, ovvero tra i 5 e 9 anni e tra i 40 e 75
anni, con l’adamantinomatoso e il papillare che si associano in maniera estremamente specifica
rispettivamente al primo e secondo picco (l’adamantinomatoso può essere riscontrato ad ogni età,
ma è più frequente nei bambini, mente il papillare si riscontra unicamente negli adulti) (Hamblin R,
2021).
Nonostante un survival rate elevato (tra l’85 e il 90% a 10 anni (Karavitaki N B. C., 2005)), questi
tumori presentano un alto tasso di recidiva post trattamento chirurgico; inoltre, i soggetti colpiti
mostrano un aumento della morbidità, un rischio di mortalità dalle 3 alle 5 volte maggiore rispetto
alla popolazione generale (non legato al tumore, quanto alle sue conseguenze) e,
complessivamente, una importante riduzione della qualità della vita (Hamblin R, 2021) (Sherlock
M, 2010). Ad incidere, in maniera preponderante, su questi aspetti sono soprattutto l’obesità, la
sindrome metabolica ed il diabete mellito, patologie che presentano tutte una aumentata incidenza
nei pazienti affetti (van Santen, 2019). Queste si estrinsecano come conseguenza del danno
ipotalamico, che si può presentare in due forme: una pre operatoria, dovuta alla compressione del
tumore a livello delle strutture ipotalamiche, ed una post operatoria, più frequente, dovuta a una
lesione diretta conseguente alla terapia chirurgica (Erfurth, 2020). Oltre a queste criticità, molti
soggetti mostrano anche altre problematiche legate alla visione, alla sfera endocrina (specie
pan/ipopituitarismo e diabete insipido centrale), alla regolazione energetica ed alla funzionalità
cognitiva e comportamentale (Zoicas F, 2012).
Alla luce di tutto ciò, l’attuale approccio al trattamento del craniofaringioma, sia questo
adamantinomatoso che papillare, non può esimersi dal prendere in considerazione il management
di queste comorbidità, sia singolarmente che in maniera congiunta, allo scopo di migliorare
l’outcome terapeutico, la prognosi, oltre che, la qualità della vita dei pazienti.
Storia
Nel 1840 von Mohr descrisse il rapido sviluppo di obesità in un caso di tumore ipofisario (Müller HL,
2019). Le sue osservazioni riguardanti gli adenomi ipofisari, assieme a quelle di Frohlic nel 1901,
portarono all’ipotesi che l’obesità adenoma-correlata fosse dovuta a un danno ipotalamico (Gao Q,
2008).
La prima descrizione di un craniofaringioma fu nel 1857 a opera del patologo tedesco Zenker, che
descrisse una massa cistica soprasellare contenente cristalli di colesterolo e gruppetti di cellule di
22aspetto simile ad epitelio squamoso a livello della pars distalis e pars tuberalis dell’ipofisi,
successivamente studiate da Lushka nel 1860. Si pensava che queste cellule fossero simili a quelle
dell’epitelio del cavo orale. Questi riscontri, assieme all’identificazione da parte di Rathke nel 1838
dell’evaginazione della sezione cefalica dell’intestino primitivo come tappa fondamentale nello
sviluppo della ghiandola pituitaria, contribuirono alla comprensione della patogenesi del
craniofaringioma, per la prima volta proposta da Mott e Barrett nel 1899 (Müller HL, 2019)
(Barkhoudarian G, 2013). Si ritiene che le manifestazioni cliniche del craniofaringioma siano state
per la prima volta descritte da Boyce e Beadles nel 1893 (Barkhoudarian G, 2013).
Nel 1902 anche Saxer descrisse un tumore composto da queste cellule; due anni dopo, poi,
Erdheim, fu il primo a descrivere in maniera accurata le caratteristiche istopatologiche del
craniofaringioma, riferendo la presenza di queste cellule unicamente nei pazienti adulti a livello
della porzione anteriore dell’infundibolo ed in gruppi od isolette di varia grandezza, forma e
numero. Poiché questi gruppi di cellule contenevano piccole cisti simili a quelle di alcuni tumori
ipofisari non ancora denominati a quel tempo, questo si convinse che entrambe le lesioni avessero
la stessa origine e li definì neoplasmi del dotto ipofisario. In maniera concorde a Mott e Barrett,
anche lui era convinto che questi tumori originassero dalle cellule squamose originanti da un dotto
craniofaringeo parzialmente involuto (Barkhoudarian G, 2013) (Über, 1904).
Nel 1909 Halstead tenterà la prima asportazione chirurgica, mentre, solo nel 1932, le cellule
descritte da Saxer sarebbero state individuate anche nei bambini da Susman (Barkhoudarian G,
2013) (Müller HL, 2019). Differenti terminologie vennero utilizzate per descrivere questo tumore,
con il termine “craniofaringioma” per la prima volta coniato da Frazier nel 1931 e successivamente
reso popolare nel 1932 da Cushing (Barkhoudarian G, 2013) (Cushing, 1932).
Epidemiologia
Il craniofaringioma è un tumore raro, con un’incidenza che va da 0,5 a 2 casi per milione di persone
ogni anno, di cui tra il 30 e 50% a presentazione infantile o adolescenziale (Garnett MR, 2007)
(Müller HL, 2019). Negli adulti rappresenta tra il 2 e il 5% dei tumori intracranici, mente nei bambini
circa il 5% di tutti i tumori, tra il 1,2 e il 4 % di tutti i tumori intracranici e il 50% dei tumori della
regione sellare e parasellare (Garnett MR, 2007) (Müller HL, 2019). Può essere diagnosticato a ogni
età, anche nel periodo prenatale e neonatale.
È stata dimostrata una distribuzione bimodale dell’età dei pazienti affetti da craniofaringioma, con
due picchi di incidenza nei bambini/adolescenti tra i 5 e 14 anni e negli adulti/anziani tra i 45 e 75
anni. (Hamblin R, 2021) (Müller HL, 2019) (Karavitaki N C. S., 2006). In maniera più specifica i
Papillari (PCPs) si ritrovano quasi esclusivamente negli adulti, con picco tra i 40 e 55 anni di età. Gli
23Puoi anche leggere

















































