Il segnale elettromiografico - Francesco ...
←
→
Trascrizione del contenuto della pagina
Se il tuo browser non visualizza correttamente la pagina, ti preghiamo di leggere il contenuto della pagina quaggiù
Università di Roma La Sapienza
Corso di Elaborazione di Dati e Segnali Biomedici 1
Facoltà di Ingegneria Civile ed Industriale
Il segnale elettromiografico
Ingegneria Clinica
A.A. 2018-2019
Francesco Infarinato, PhD
Laboratorio di Bioingegneria della Riabilitazione
IRCCS San Raffaele Pisana
www.sanraffaele.it
francesco.infarinato@uniroma1.it
www.infarinato.it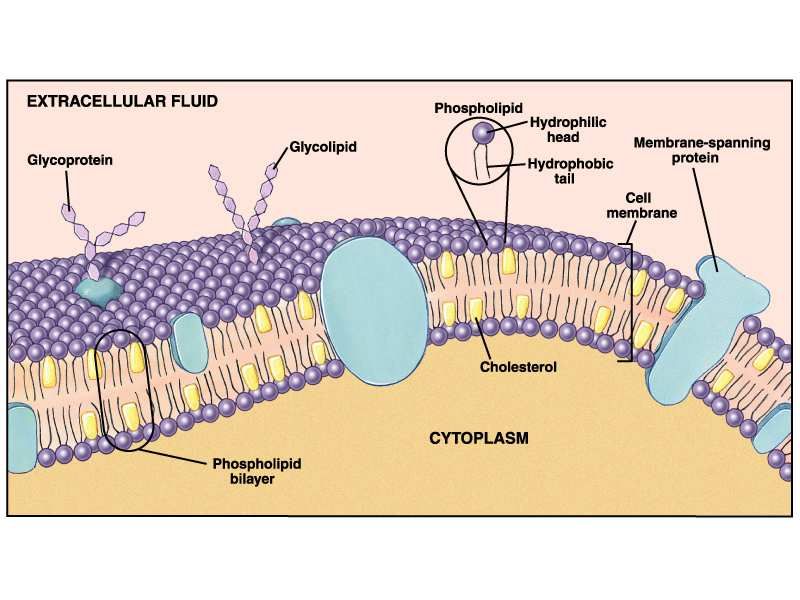
Sommario
• Richiami di anatomia e fisiologia muscolare
• Segnale elettrico muscolare
• Elettromiografia ad ago e di superficie
• Parametri globali del segnale EMG
• Fenomeni di fatica mioelettrica
• Velocità di conduzione media delle fibre
muscolari
2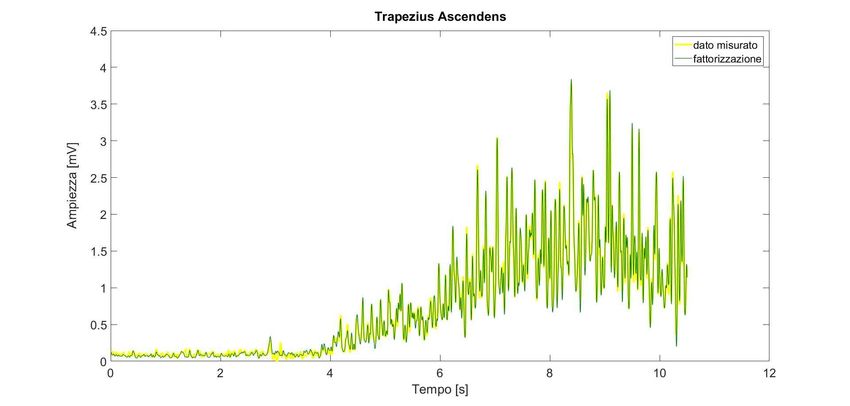
Richiami di anatomia e fisiologia
• Cellule eccitabili
• Potenziale a riposo e potenziale di azione
• Struttura delle fibre muscolari
– Fisiologia della contrazione muscolare
– Motoneuroni – unità motorie
– Strategie di modulazione della forza
• Principio del reclutamento
• Fenomeni di fatica muscolare
3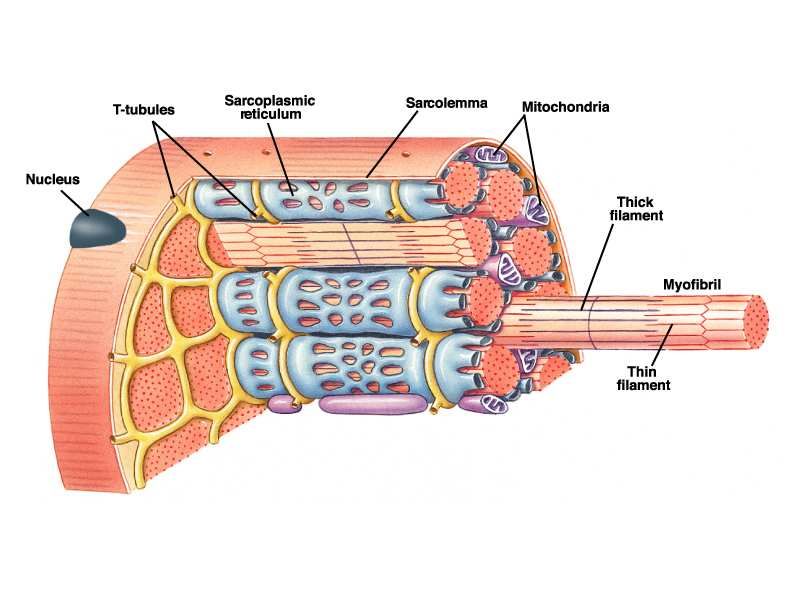
Membrana Cellulare
• Composta di molecole fosfolipidiche e proteine
• Presenta canali di ingresso-uscita
4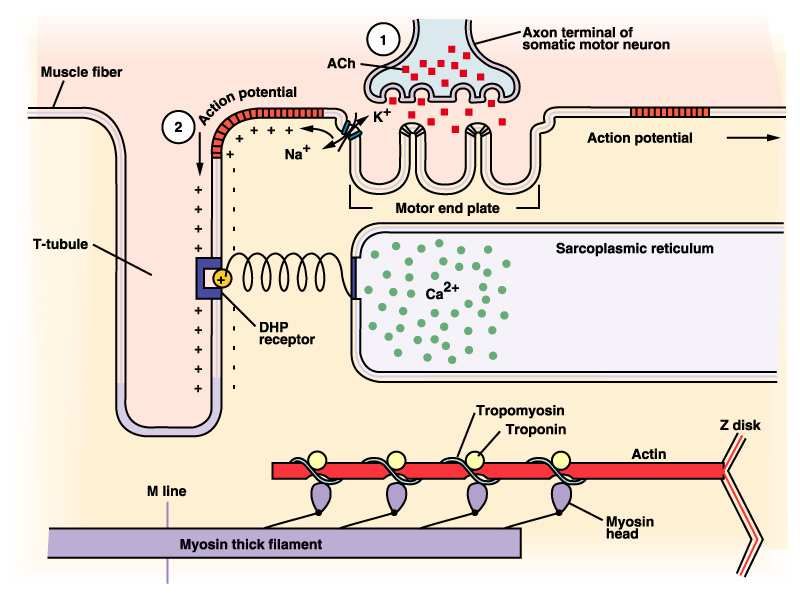
Equazione di NERNST
RT [ K +]
Vi = ln ext
zF [ K +] int
Equazione di Goldman-Hodgkin-Katz
RT P[K +] + P [ Na+] + P [Cl−]
Vm = ln k ext Na ext Cl int
F P[K +] + P [ Na+] + P [Cl−]
k int Na int Cl ext
R=Costante universale degi GAS (8314.9 J/Kg.Mol.K)
Z=valenza dello ione
T=Temperatura assoluta °K
F=costante di Faraday
Pi=permeabilità della membrana cellulare allo ione I
[]ext=concentrazione esterna
5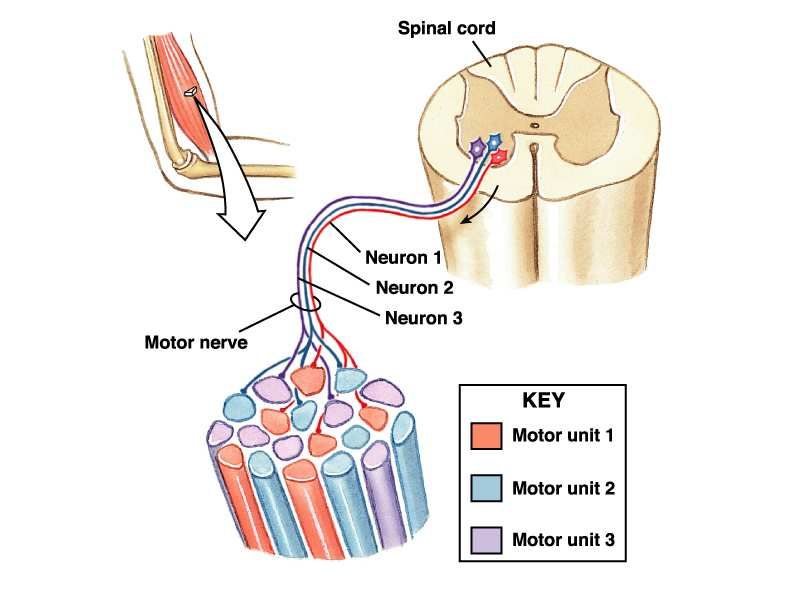
Cellule eccitabili
In condizione di non eccitazione, il
canale è CHIUSO
• Sono le cellule che
posseggono canali
voltaggio/chemio
dipendenti all’interno
della loro membrana
Un impulso elettrico o chimico causa
l’apertura del canale, consentendo il
passaggio di ioni attraverso la membrana
6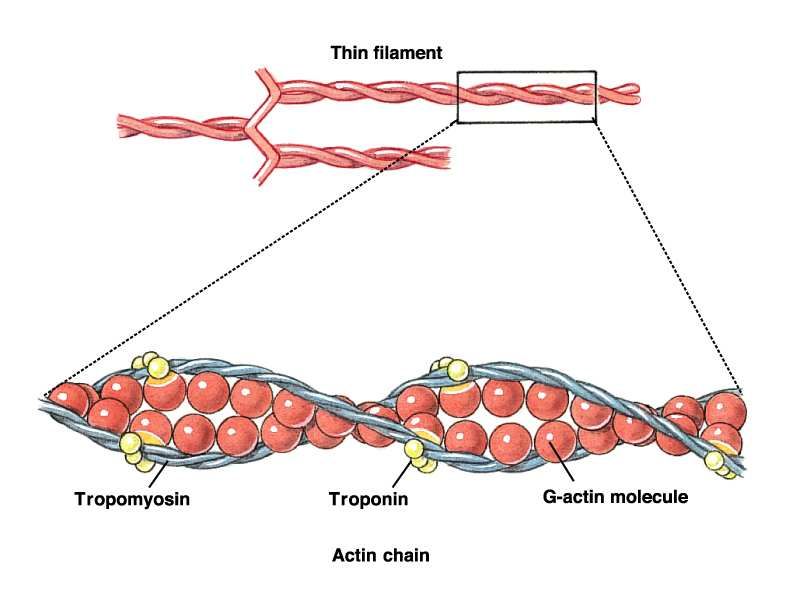
• Lo stato dei canali voltaggio-dipendenti
influenza la permeabilità della cellula al
passaggio di alcuni ioni.
La cellula eccitabile presenta un potenziale non
nullo se misurato rispetto all’esterno
La variazione di permeabilità comporta un trasporto di
cariche al passare del tempo, ovvero una corrente.
La perturbazione dello spazio causata da questo flusso di
corrente può essere misurata.
7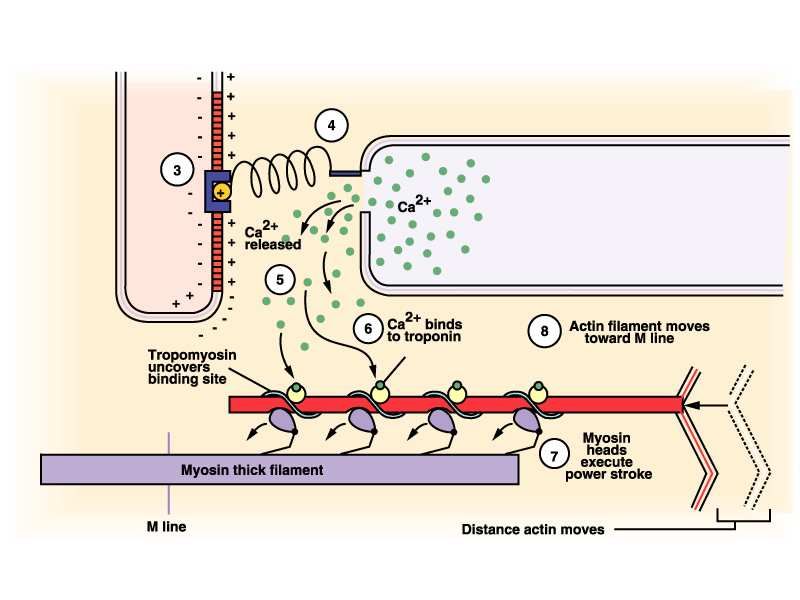
Potenziale di Azione
• Uno stimolo chimico interessa una porzione della
membrana
• L’effetto si propaga nelle porzioni limitrofe dando
luogo a una progressiva depolarizzazione dell’intera
membrana (conduzione continua).
8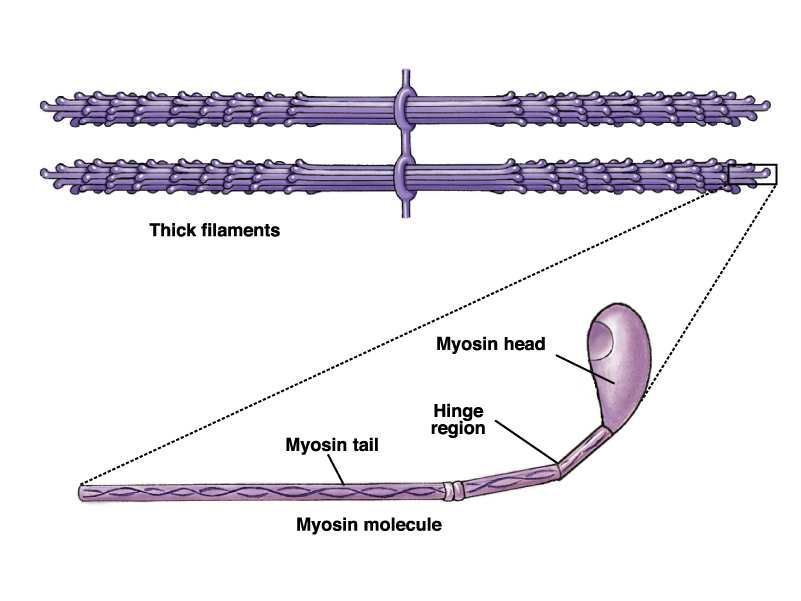
Muscolatura Scheletrica
• I muscoli scheletrici sono
responsabili del
movimento volontario
• Rappresentano il 45% del
tessuto muscolare
complessivo presente
nell’organismo
• Sono collegate al SNC
tramite le giunzioni
neuromuscolari
(Innervation Zones, IZ)
9
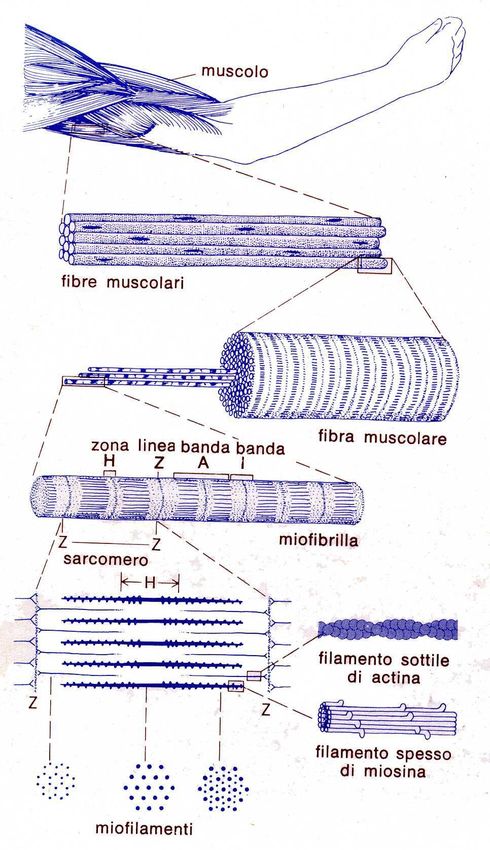
Anatomia delle fibre muscolari
•Ventre muscolare
•Fibra muscolare
•Miofibrilla
•Sarcomero
•Proteine contrattili e di
controllo
10Anatomia della fibra muscolare
11Sarcomero
• È l’unità funzionale atomica del movimento
• Contiene le proteine contrattili e quelle di controllo
12Proteine del sarcomero
• Miosina-> filamento spesso
• Actina->filamento sottile
} Contrattili
•Troponina
•Tropomiosina
}Di controllo
13Fisiologia della contrazione muscolare
• Teoria dei filamenti scorrevoli
– La contrazione avviene per lo scorrimento delle
molecole di miosina su quelle di actina grazie
all’energia fornita dall’ATP (adenosin tri fosfato)
14Teoria dei filamenti scorrevoli 1/3
In condizioni di riposo le cisterne del
Calcio sono chiuse e actina e miosina
non possono legarsi
Quando arriva un potenziale di
Azione le cisterne si aprono, il Ca2+ si
lega alla troponina liberando il sito
attivo dell’actina su cui si attacca la
testa della miosina.
15Teoria dei filamenti scorrevoli 2/3
…Il legame tra calcio e troponina causa una
modificazione nella forma della tropomiosina
che consente la formazione del legame tra
Actina e miosina
Una molecola di ATP legata alla testa
della miosina fornisce l’energia
necessaria alla rotazione della
stessa, facendo perno sul filamento sottile
e liberando un fosfato inorganico
16Teoria dei filamenti scorrevoli 3/3
L’ADP viene rilasciato e una nuova
molecola di ATP si lega alla miosina; la
testa della miosina si stacca dall’actina
La molecola di ATP appena legata
viene scissa in ADP+P e il sistema è
pronto per un nuovo ciclo.
Se nel sarcoplasma è presente ancora
Ca2+ la testa della miosina compie
un’altra traslazione sulla molecola di
actina
1718
A monte…
• Impulso dal SNC
• Rilascio di acetilcolina a livello della placca di
innervazione
• Apertura delle cisterne del calcio
• Scorrimento delle proteine contrattili
CONTRAZIONE
19Propagazione del PDA
• La depolarizzazione di una porzione del
sarcolemma causa una variazione di
permeabilità agli ioni delle porzioni ad esso
limitrofe, dando vita a un fenomeno di
conduzione continua
20Modellizzazione del potenziale di azione
Il PDA muscolare si propaga lungo il sarcolemma come un tripolo elettrico
lungo una linea
www.lisin.polito.it
21Tipi di fibre muscolari
• Diverse isoforme della catena pesante della
miosina (MHC)
• Diverse proprietà metaboliche, meccaniche e di
resistenza
Le proprietà dei diversi tipi di fibre muscolari, sono legate anche al motoneurone che le
innerva: è quindi corretto parlare anche di motoneuroni di tipo I e II
22In particolare…
Maggiore quantità di mitocondri,
maggiore vascolarizzazione->fibre
“rosse”
Tipo I (slow twitch Tipo II (fast twitch
ST) FT)
Metabolismo ossidativo glicolitico
Dimensione piccola grande
Segnale di ampiezza inferiore
Velocità di contrazione bassa alta
Adatta a movimenti lenti e di
Resistenza alta lunga durata bassa
all’affaticamento
Velocità di conduzione bassa alta
23Unità motoria
• L’insieme di un motoneurone e delle fibre da esso innervate
Ogni motoneurone innerva più fibre, ma ogni fibra può essere innervata da un solo
motoneurone
24Rapporto α-motoneurone/numero di fibre
• Il numero e il tipo di fibre muscolari di una unità
motoria in un muscolo ci danno indicazioni sulla sua
funzione
Muscoli oculomotori: 10 Quadricipite femorale:
fibre per UM (regolazione 10000 fibre per UM
fine del movimento (postura eretta, corsa,
dell’occhio..) camminata..)
25Sistemi di modulazione della forza
• Principio del reclutamento
• Regolazione della frequenza di scarica
• Attivazione coordinata di muscoli agonisti e
antagonisti
• Sinergie di gruppi muscolari
26Principio del reclutamento (size principle)
[Henneman 1977]
• Durante una contrazione il numero delle unità motorie attive
cresce al crescere della forza erogata e il reclutamento procede
ordinatamente dalle unità motorie più lente (e piccole) a quelle
più veloci (e grandi)
27Frequenza di scarica
• La frequenza di scarica delle
UM aumenta all’aumentare
della forza
• Da 80%MVC l’incremento di
forza è dovuto
ESCLUSIVAMENTE
all’aumento della frequenza di
scarica (il reclutamento è già
completo)
28Fatica
• Aumento dei prodotti catabolici
– Anidride carbonica (CO2)
– Ioni H+
• Diminuzione di ossigeno in seguito a
modificazioni geometriche del muscolo e dei
vasi che lo irrorano
• Diminuzione dei substrati energetici
– riduzione glicogeno muscolare
29L’elettromiografia – un po’ di storia
1790 – Galvani e Volta mostrano gli effetti della contrazione indotta
1840 – Carlo Matteucci descrive gli effetti della corrente sugli animali
1860 – Emil Heinrich Du Bois-Reymond: il cervello produce corrente
1862 – Guillaume Duchenne sperimenta la stimolazione muscolare
30L’elettromiografia – un po’ di storia
1790 – Galvani e Volta mostrano gli effetti della contrazione indotta
1840 – Carlo Matteucci descrive gli effetti della corrente sugli animali
1860 – Emil Heinrich Du Bois-Reymond: il cervello produce corrente
1862 – Guillaume Duchenne sperimenta la stimolazione muscolare
1929 – Charles Scott Sherrington definisce le unità motorie
1942 – Herbert Jasper costruisce il primo elettromiografoSegnale elettromiografico
• Generazione del segnale (somma di MUAPT)
• Caratteristiche del segnale EMG
– Contenuto in frequenza
– EMG e unità motorie
– Segnale da singola fibra, volontario e stimolato
– Intervalli di stazionarietà
32Generazione del segnale EMG
Rappresenta la somma dei contributi dati dall’attività ASINCRONA delle unità
Motorie reclutate in un certo istante di tempo
33Contributo delle UM al segnale sEMG
• Dimensione delle fibre attive
• Quantità di fibre per UM
• Distanza dal punto di prelievo
• Presenza di disomogeneità del tessuto
interposto
34Caratteristiche del segnale EMG
Ampiezza 50µV÷5mV
Contenuto in frequenza 10÷450Hz
Intervallo di stazionarietà 500÷1000ms in condizioni
isometriche
35Elettromiografia
• Sensori per il prelievo del segnale (di profondità e di
superficie)
• Modalità di prelievo
– Monopolare
– Singolo differenziale
– Doppio differenziale
• Elettromiografo
• Conversione analogico-digitale
– Contenuto in frequenza ricostruzione del segnale
• Differenze fra segnale intramuscolare e di superficie
36Elettrodi per EMG
Needle EMG electrode
PRO CONTRO
Segnale meno Tecnica invasiva
rumoroso
Maggiore Posizionamento
contenuto nelle alte elettrodo doloroso e
frequenze scomodo
Mostra le proprietà locali del muscolo in
esame
37Elettrodi per EMG
PRO CONTRO
Facile Segnale filtrato dal Flottante
applicazione e grasso subcutaneo
riposizionamento
Non invasiva Risente
Attivo
maggiormente
(amplificatore
degli artefatti da
on board)
movimento
Fornisce informazioni globali sul
muscolo in esame
Schiera lineare
38Elettromiografo
Accoppiamento
optoelettronico o induttivo
Seconda amplificazione
Elettrodo
Blocco di
filtraggio
Prima
amplificazione
CPU
M A/D
U Converter
RAM
X
HDD
39Modalità di prelievo del segnale EMG
-1
+1 Monopolare
1
-1
+1 -1
DD1
+1
-1
Doppio differenziale +1
SD
-1
DD2
+1
-1
+1
-1
SD Singolo differenziale
+1
40Condizionamento del segnale: filtraggio
Notch
HighPass (10Hz)
(band-stop 49÷51Hz)
LowPass (500Hz)
41Parametri globali del segnale EMG
• Grandezze caratteristiche del segnale EMG
– RMS, ARV, MDF, MNF: tipi di fibre e fatica
mioelettrica
– Velocità di conduzione media delle fibre muscolari
– RQA %DET, sincronizzazione e determinismo
42Parametri nel tempo
Average Rectified Value
T /2 N 2
1 1
ARV = ∫ | v(t ) | dt
T −T / 2 N
∑| x
n=− N 2
n |
43Parametri nel tempo
Root Mean Square
T /2 N 2
1 1
RMS = ∫ ∑ n
2 2
v (t )dt x
T −T / 2 N n=− N 2
44Parametri in frequenza
∞
∫ P ( f ) df = M
0
0
MDF
1
Median Frequency MDF: ∫ P ( f )df = M 0
0 2
∞ ∞
Mean Frequency MNF= ∫ fP( f )df / ∫ P( f )df = M / M
0 0
1 0
45Studio dell’affaticamento mioelettrico attraverso la MDF
Contrazione Isometrica BB 80%MVC
sEMG
t0=4s t [s] f [Hz]
MDF0
Lo spostamento della MDF
misura il cambiamento del
contributo oscillatorio delle
fibre e quindi è indice delle
dinamiche di reclutamento
t1=22.5 s t [s] f [Hz]
MDF1 46Studio dell’affaticamento mioelettrico attraverso la MDF
Hz
Forza (unità arbitrarie) 80%MVC
isometrica
MDF
t [s]
47Il concetto di velocità di conduzione (CV)
10 mm
Nota la distanza tra due punti di prelievo, è
possibile stabilire la velocità di 2
propagazione del segnale elettromiografico
noto il tempo necessario a passare da un MUAP PROPAGATION
1
punto di prelievo all’altro
Canali EMG
0
s
V =
∆t = 2ms
-1
-1
MFCV = 5ms
sEMG Amplitude [mV]
-2
t
0.114 0.116 0.118 0.120 0.122 0.124 0.126 0.128 0.130 0.132
Time [s]
EMG Ch1
EMG Ch 2
Tempo
48Il concetto di velocità di conduzione (CV)
Canali EMG
∆s
∆t Tempo 49Problematiche aperte
• Perché stimare la CV?
• Come stabilire con (ragionevole) certezza se
l’evento osservato nei due punti è lo stesso?
• Quali sono le principali strategie
implementative per stimare la CV?
• CV e configurazioni elettrodiche
50Il ciclo del passo (Stride)
Un ciclo di passo è la sequenza di eventi durante la
locomozione definita da quando un piede tocca il terreno
fino a quando lo stesso piede non tocca il terreno di nuovo.
Il passo implica la propulsione del centro di gravità e può
esseere suddiviso in due fasi principali:
Stance: Il periodo in cui il piede è a contatto col suolo (0-60% stride)
Swing: Il periodo in cui il piede è sollevato dal suolo per permetteree
l’avanzamento dell’arto (60-100% stride)
51Correlati fisiologici del passo – Regolazione sovraspinale
Regioni mesencefaliche: gestiscono attivazione e velocità
Cervelletto: Responabile dell’accuratezza del gesto
Corteccia somatosensoriale: Processa l’informazione visiva
Afferenze somatosensoriali esterocettive e propriocettive:
permettono l’adattamento di pattern motori stereotipati all’ambiente
52Correlati fisiologici del passo – Generatori centrali di
pattern
I generatori di pattern centrali sono circuiti neuronali che, quando
attivati, possono produrre pattern motori ritmici come camminare,
respirare, volare e nuotare in assenza di input sensoriali o discendenti
che portino informazioni di temporizzazione specifiche.
Nel caso specifico del cammino umano, tali circuiti scaricano
ritmicamente nel midollo spinale agendo da driver del comportmento
locomotorio e producendo il ritmo di base del passo.
La connettività effettiva tra i sottogruppi di interneuroni e singoli
pool di neuroni motori che innescano la contrazione di muscoli
particolari è ancora sotto indagine.
53Le sinergie muscolari
Sinergìa s. f. [dal gr. der. di συνέργω «cooperare» (comp. di σύν «con,
insieme» e ἔργω «operare, agire»)]. – Azione combinata e
contemporanea, collaborazione, cooperazione di più elementi in una
stessa attività, o per il raggiungimento di uno stesso scopo o risultato,
che comporta un rendimento maggiore di quello ottenuto dai varî
elementi separati […]
Il SNC, generando comandi motori, attiva una combinazione lineare
di sinergie muscolari, ognuna delle quali attiva un insieme di muscoli,
secondo pesi diversi, come un’unità singola.
La varietà del repertorio motorio viene quindi prodotta dalla
modulazione, a livello corticale e dei circuiti riflessi, di segnali che
selezionano, attivano e combinano in maniera flessibile poche
strutture neurali fisse a livello spinale e del tronco encefalico.
54Le sinergie muscolari
La sinergia muscolare è quindi il meccanismo per mezzo del quale
più gruppi muscolari concorrono simultaneamente ed in maniera
coordinata.
Numerosi studi sulla fisiologia del movimento nei vertebrati hanno
portato a considerazioni in merito all’esistenza di rudimentali moduli
di attivazione muscolare che hanno origine nella colonna vertebrale e
che si combinano tra loro per dare luogo al movimento.
Non è attualmente ancora chiaro il meccanismo attraverso cui le aree
corticali responsabili del movimento operino attraverso questi moduli
per generare il movimento globale ma è rilevante l’importanza dello
studio di come questi moduli operino per la comprensione delle
sinergie muscolari.
55Le sinergie muscolari
Fondamentale per lo studio delle sinergie muscolari è lo sviluppo di
un algoritmo in grado di costruire una connessione tra la fisiologia del
movimento umano e i segnali bioelettrici prelevati sulla cute del
soggetto sperimentale in corrispondenza delle principali fasce
muscolari coinvolte nel movimento stesso.
L’algoritmo per valutare le sinergie muscolari si basa sullo strumento
di Fattorizzazione Matriciale Non Negativa (NNMF) il quale
permette di passare da una matrice complessa X che rappresenta le
acquisizioni EMG (n canali x m campioni) al prodotto di due matrici
di complessità inferiore W e H, a meno di un errore E:
56Le sinergie muscolari
L’obiettivo dell’analisi è minimizzare l’errore E attraverso la scelta
delle matrici W e H.
La colonne di W rappresenteranno i k moduli che descrivono le
sinergie degli n muscoli coinvolti nell’acquisizione.
Le righe di H rappresenteranno i k coefficienti di attivazione composti
da m campioni.
Dopo aver stabilito le caratteristiche dell’algoritmo per la
minimizzazione dell’errore e per la scelta di k potrò ricostruire il
segnale EMG (la Matrice X) attraverso il prodotto tra Sinergie e
Coefficienti di attivazione
57Le sinergie muscolari
Il set di segnali EMG subisce una profonda pre-elaborazione le cui
fasi fondamentali possono essere così riassunte:
1. Filtraggio passa-alto (50Hz)
2. Rettificazione
3. Filtraggio passa-basso (20Hz)
4. Normalizzazione
5. Smoothing (mooving average)
Queste operazioni incideranno sull’aspetto del segnale nel tempo e sul
suo spettro di densità di potenza.
58Le sinergie muscolari
59Le sinergie muscolari Le prove sperimentali suggeriscono che il SNC possa aggirare le complessità computazionali del controllo motorio generando comandi attraverso una combinazione lineare di sinergie motorie, ciascuna in grado di attivare un gruppo di muscoli come una singola unità. La scelta del numero K di sinergie è dunque fondamentale e non può prescindere dalla qualità della ricostruzione operata dalla NNMF. Per garantire una ricostruzione ottimale del segnale registrato si procede dunque con la classificazione della bontà dell’algoritmo in base al valore R2 della ricostruzione. •yi = dati stimati •xi = dati osservati •x = media dei dati osservati R2 o coefficiente di determinazione (rapporto tra la devianza spiegata e la devianza totale) è una proporzione tra la variabilità dei dati e la correttezza del modello statistico utilizzato e stabilisce la qualità della predizione effettuata su una sequenza di valori misurati 60
Le sinergie muscolari
La scelta del numero k di sinergie viene fatta considerando il minor numero corrispondente
ad una buona approssimazione del dato misurato (con R2>0.8); allo stesso modo si può
interpretare l’azione del sistema nervoso come un controllore in grado di ottimizzare i
processi e ridurne il più possibile la spesa energetica.
61Puoi anche leggere

















































