Herpesviridae AA 2019-2020 - G. Di Bonaventura Università di Chieti-Pescara CI Microbiologia e Microbiologia Clinica - Scuola di Medicina e ...
←
→
Trascrizione del contenuto della pagina
Se il tuo browser non visualizza correttamente la pagina, ti preghiamo di leggere il contenuto della pagina quaggiù
Herpesviridae
G. Di Bonaventura
Università di Chieti-Pescara
CI Microbiologia e Microbiologia Clinica
AA 2019-2020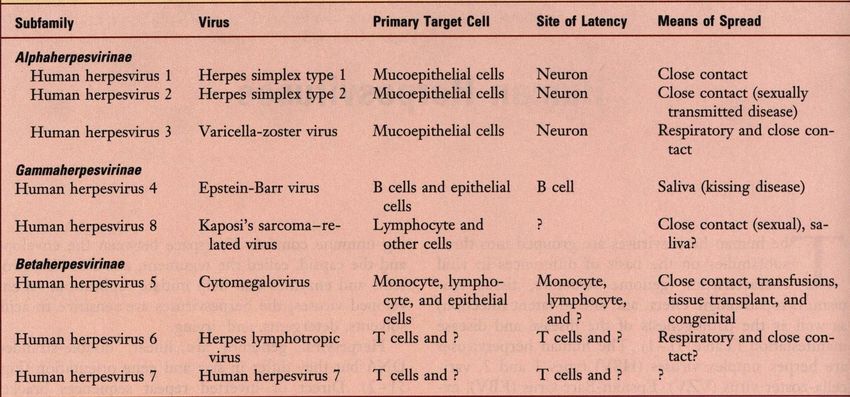
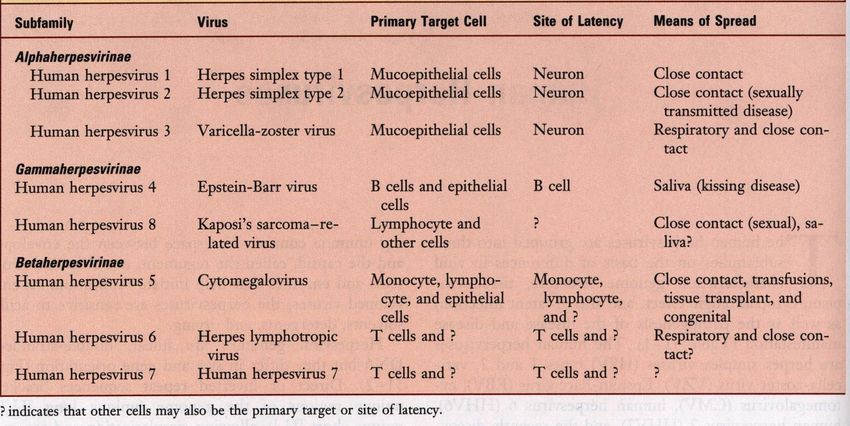
Proprietà caratteristiche degli herpesvirus umani
Medical Microbiology, 4th ed., Murray, Rosenthal, Kobayashi & Pfaller, Mosby Inc., 2002, Table 51-1.
Alphaherpesvirinae: ampio spettro d’ospite (inoculazione per scarificazione o per via
intracerebrale), si replicano in diversi tipi di colture cellulari con caratteristico effetto citopatico e
formazione di inclusioni nucleari.
Gammaherpesvirinae: specie-specifici, si replicano solo in colture cellulari della specie sensibile.
Betaherpesvirinae: ristretto spettro d’ospite, tropismo per le cellule linfoidi.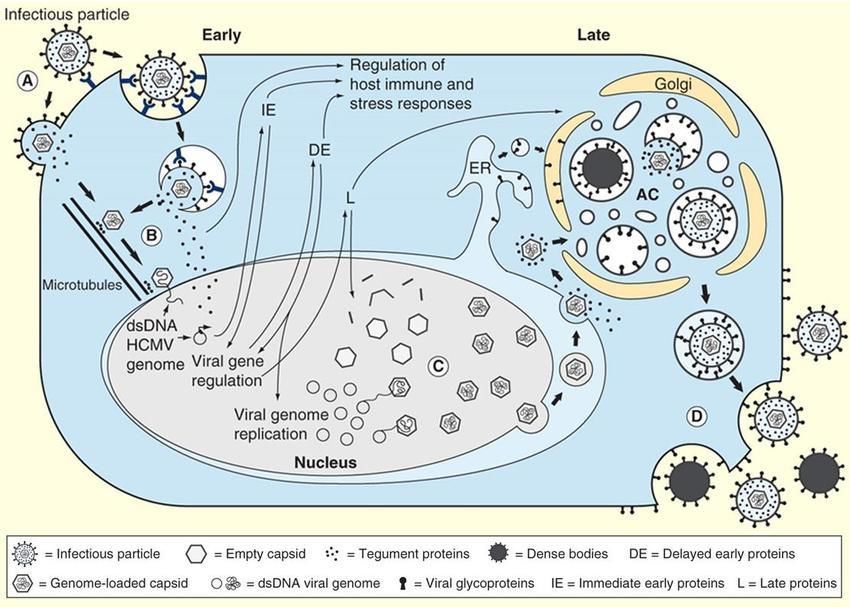
Herpesvirus: genoma
The genomes of the herpesvirus are doubled-stranded DNA. The length and complexity of the genome differ for
each virus. Inverted repeats in herpes simplex virus (HSV), varicella-zoster virus (VZV), and cytomegalovirus (CMV)
allow the genome to recombine with itself to form isomers. Large genetic repeat sequences are boxed. The
genomes of HSV and CMV have two sections, the unique long (UL) and the unique short (US), each of which is
bracketed by two sets of inverted repeats of DNA. The inverted repeats facilitate the replication of the genome but
also allow the UL and US regions to invert independently of each other to yield four different genomic configurations,
or isomers. VZV has only one set of inverted repeats and can form two isomers. Epstein-Barr virus (EBV) exists in
only one configuration, with several unique regions surrounded by direct repeats. Purple indicates direct repeat DNA
sequences; green indicates inverted repeated DNA sequences. HHV6: human herpesvirus 6.
[Medical Microbiology, 4th ed., Murray, Rosenthal, Kobayashi & Pfaller, Mosby Inc., 2002, Table 51-2]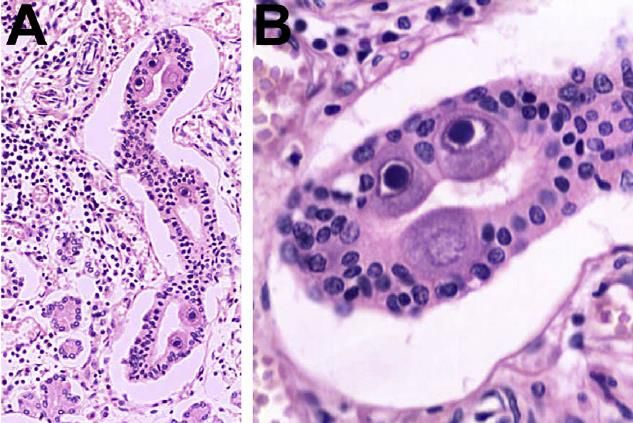
Herpesvirus: ciclo replicativo
1. Virion binds to the cell surface, and then envelope fuses with the plasma membrane releasing the capsid
and tegument proteins into the cytoplasm. The vhs protein causes degradation of mRNAs (shut-off).
2. Capsid is transported to the nuclear pore, where the viral DNA is released into the nucleus.
3. Viral DNA circularizes and is transcribed by host RNA polymerase II to give IE (immediate early, 2-4 h
post-infection) or alpha mRNAs. IE gene transcription is stimulated by the VP16 tegument protein.
4. Five of the six IE proteins regulate viral gene expression in the nucleus, and transactivate E (early, 5-8 h
p.i.) or beta gene transcription. E proteins are involved in replicating (rolling circle) the viral DNA.
5. Viral DNA synthesis stimulates L (late, 9 h p.i.) or gamma gene expression. The L proteins are
involved in assembling the capsid in the nucleus and modifying the membranes for virion formation.
6. The filled capsid buds through inner membrane to form an enveloped virion that exits from the cell.
[Fields Virology, 4th ed, Knipe & Howley, eds, Lippincott Williams & Wilkins, 2001]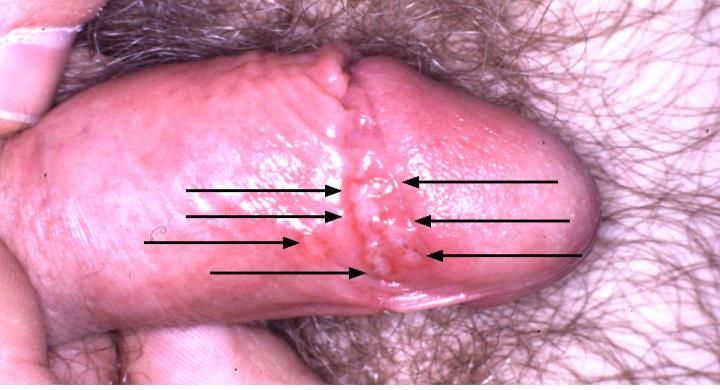
Herpesvirus: “maturazione” degli involucri virali
HSV capsids bud through the inner nuclear membrane, forming an enveloped virion particle.
Egress of the virions from the host cell may occur by either of two general pathways.
A: The (primary) envelope fuses with the outer nuclear membrane, de-enveloping the capsid and
releasing it into the cytoplasm. The capsid then buds into the Golgi apparatus, forming an enveloped
(secondary enevelope) virion, which is transported to the surface by vesicular transport.
B: The virion particle buds through the outer nuclear membrane and is transported by vesicular
movement through the Golgi apparatus to the exterior of the cell.
[Fields Virology, 4th ed, Knipe & Howley, eds, Lippincott Williams & Wilkins, 2001]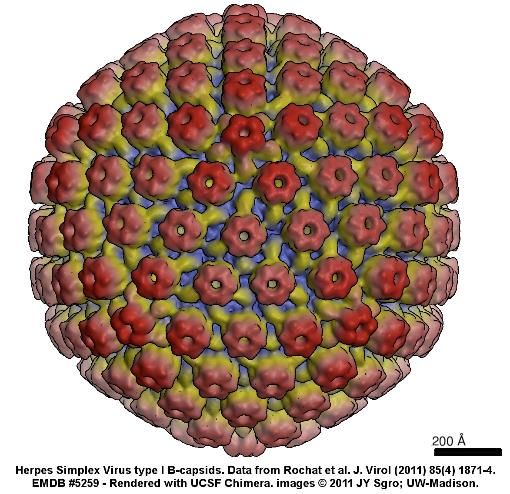
Alpha-Herpesvirinae ▪ Spettro d’ospite più ampio fra tutti i membri della famiglia. ▪ In vitro si propagano facilmente in molti tipi cellulari. ▪ Ciclo riproduttivo piuttosto breve. ▪ Nell’ospite naturale infezione LITICA soprattutto in cellule epiteliali (epidermide e mucosa), infezione LATENTE in cellule neuronali (neurotropismo alfa). ▪ Portano alla completa distruzione delle cellule infettate (citolisi). ▪ Nell’uomo: Herpes simplex virus (HSV-1 e HSV-2), Virus Varicella Zoster (VZV) ▪ Negli animali: Herpesvirus bovini (BoHV-1 e BoHV-2), Herpesvirus equini (EHV-1 e EHV-4)
Herpesvirus simplex
Struttura
Virione: sferoidale, d: 200 nm
Peplos di origine cellulare, contiene almeno 11
glicoproteine tra cui:
▪ gB, gC, gD, gH, gE, gI: «concentrano» i virioni
alla superficie della cellula sensibile legandoli
agli eparan-solfati
▪ gD funge da antirecettore, riconoscendo
HVEM (HerpesVirus-Entry-Mediator, recettore
per TNF-), nectina-1 e -2 (Ig ICAM)
▪ gB: fase di adsorbimento e fusione
▪ gC, gE, gI: proteggono i virioni dalla immunità
umorale (legando Complemento e Fc di Ig)
Tegumento (matrice): proteine virus-specifiche
Capsìde: icosaedrico, 100 nm, 162 capsomeri
Nucleoide: struttura a rocchetto con avvolto il DNA
Genoma: DNA ds (150 kbp), lineare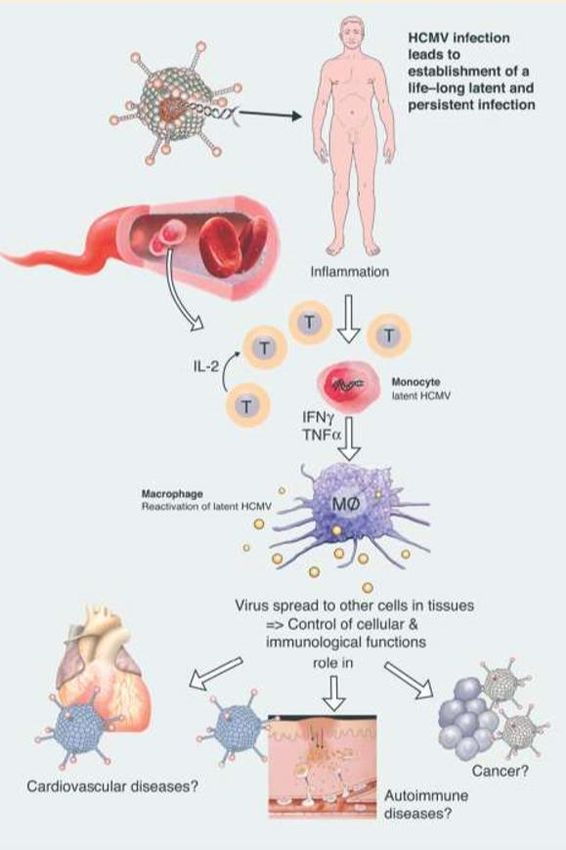
Herpesvirus: genoma The genomes of the herpesvirus are doubled-stranded DNA. The length and complexity of the genome differ for each virus. Inverted repeats in herpes simplex virus (HSV), varicella-zoster virus (VZV), and cytomegalovirus (CMV) allow the genome to recombine with itself to form isomers. Large genetic repeat sequences are boxed. The genomes of HSV and CMV have two sections, the unique long (UL ) and the unique short (US ), each of which is bracketed by two sets of inverted repeats of DNA. The inverted repeats facilitate the replication of the genome but also allow the UL and US regions to invert independently of each other to yield four different genomic configurations, or isomers. VZV has only one set of inverted repeats and can form two isomers. Epstein-Barr virus (EBV) exists in only one configuration, with several unique regions surrounded by direct repeats. Purple indicates direct repeat DNA sequences; green indicates inverted repeated DNA sequences. HHV6=human herpesvirus 6. [Medical Microbiology, 4th ed., Murray, Rosenthal, Kobayashi & Pfaller, Mosby Inc., 2002, Table 51-2]
Herpes-Simplex Virus Ciclo biologico: Latenza HSV replica nella sede di infezione primaria (cute), quindi migra - per via nervosa (o più raramente per via ematica) - verso i gangli nervosi dorsali sensitivi (HSV-1: gangli trigeminali; HSV-2: gangli sacrali). In sede gangliare il virus entra in una fase di latenza: il DNA virale si trova nel neurone in forma circolare ed episomale e non viene trascritto, ad eccezione del LAT (Latency- Associated Transcript) che nel nucleo trascrive per mi(micro)RNA inibenti l’espressione di proteine necessarie per il ciclo replicativo virale. Il mantenimento della latenza è regolato dalla immunità cellulo-mediata. In presenza di opportuni stimoli (traumi in sedi innervate da neuroni sensitivi infetti; stress emotivi, fisici od ormonali), l’infezione si “riattiva” con replicazione virale e progressione verso la sede di infezione primaria. Frequenza (25% circa degli infetti presenta riattivazioni) e gravità delle “riattivazioni” dipendono dal numero di neuroni infettati e da immunocompetenza (asintomatica negli immunocompetenti, infezione disseminata negli immunocompromessi). I virus “riattivati” infettano altri neuroni, consentendo la persistenza dell’infezione (lifelong).
Herpes-Simplex Virus
Latenza: meccanismo di regolazione
Il gene ICP4 codifica un importante transattivatore dei geni associati alla infezione litica-
produttiva.
Nei neuroni, il legame di proteine NRSF (Neuronal Restrictive Silencing Factor) e REST
(Repressor Element Silencing Transcription Factor) a ICP4 provoca la ipoacetilazione del
promotore di ICP4 che reprime la trascrizione di questo gene e, pertanto, anche la
trascrizione degli altri geni litici.
Pinnoji et al, Virology Journal 2007
Di contro, la proteina ICP22 può causare la dissociazione di NRSF dal gene ICP4,
sbloccando il silenziamento del DNA virale.
Il virus può quindi essere riattivato dalla latenza mediante stress che sbloccano il
silenziamento del DNA virale: malattie come raffreddore, influenza, eczema, o stress
emozionali e fisici, esposizione ai raggi UV, affaticamento, traumi.Herpes-Simplex Virus
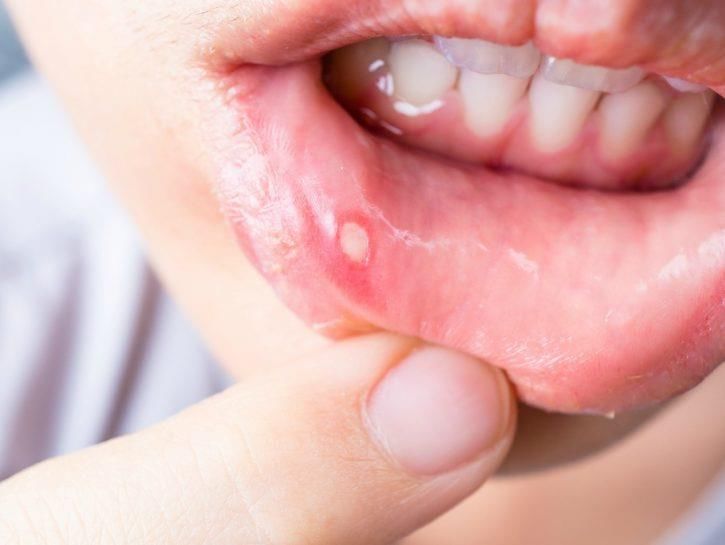
HSV-1: malattia
Contagio interumano (soprattutto nella prima infanzia): saliva, posate, bicchieri.
Causa lesioni vescicolari confluenti (a “grappolo”) a localizzazione periorale cutanea (erpete
labiale) e/o nella mucosa orale (gengivo-stomatite).
L’infezione può essere asintomatica.
Occasionalmente, HSV-1 può infettare la congiuntiva e la cornea (autoinoculazione),
causando cheratocongiuntivite con compromissione della funzionalità visiva.
Raramente, diffonde all’encefalo, causando encefalite necrotico-emorragica.
Può causare lesioni a livello genitale (il sito di infezione non è predittivo del tipo di HSV).Herpes-Simplex Virus HSV-2: malattia Contagio interumano (età adulta): via sessuale. Il virus penetra la cute/mucosa attraverso lesioni e si replica nella sede di penetrazione iniziale. Le lesioni cutanee/mucose hanno caratteristiche similari a quelle causate da HSV-1. L’infezione può essere asintomatica. Il contagio del neonato attraverso il canale del parto può causare infezioni erpetiche disseminate (infezioni oculari, encefaliti, meningiti). Mortalità >80% (se non trattate) e comunque gravi sequele neurologiche anche a seguito di guarigione. Può causare lesioni nella regione periorale/orale (sito di infezione non è predittivo del tipo di HSV).
Herpes-Simplex Virus Diagnosi Isolamento virale in colture cellulari a partire da: liquido delle vescicole muco- cutanee, materiale da raschiamento di ulcere corneali, liquor. Nell’esame citodiagnostico di Tzanck il materiale - raccolto per raschiamento della base di vescicola/bolla aperta di recente/ulcere corneali - viene colorato con Giemsa, per la ricerca delle tipiche cellule giganti polinucleate ed i cheratinociti atipici (acantolitici con grandi nuclei). Caratteristico effetto citopatico policariogenico: presenza di cellule multinucleate giganti con corpi inclusi intranucleari (corpi di Cowdry di tipo A).
Herpes-Simplex Virus
Diagnosi
L’identificazione definitiva avviene mediante dimostrazione di:
▪ antigeni virali mediante IF (impiego di Abs monoclonali anti-HSV1 e anti-HSV2)
direttamente sul materiale patologico o in colture infettate con il campione clinico
▪ dimostrazione del DNA virale mediante PCR
La ricerca sierologica è priva di significato per HSV-1, sia per l’elevato livello di Abs in
pazienti asintomatici che per la diffusione dell’infezione nella popolazione
(sieroprevalenza: >90%). Ricerca di Ig anti-gpD per HSV-2.Herpes-Simplex Virus Terapia Aciclovir (deossiguanosina aciclica) Inibisce la replicazione del DNA virale agendo da analogo strutturale dei nucleosidi. Meccanismo di azione: 1. aciclovir viene fosforilato da enzimi virali 2. l’analogo trifosfato presenta maggiore affinità (vs. nucleotidi «normali») per il sito attivo di DNA-pol, così inibisce la trascrizione fungendo da «terminatore della sintesi» Efficace per: - riduzione durata della manifestazione clinica - dilatare l’intervallo temporale tra episodi ricorrenti Si associa a scarsi effetti collaterali in quanto dotato di elevata selettività
Herpes-Simplex Virus Controllo Scarsa efficacia di vaccini a virus formati da virus vivi ma attenuati, oppure da virus privati del DNA. Attualmente in sperimentazione vaccini contenenti gD purificata da colture cellulari infette od a seguito di clonaggio. Limitata protezione vaccinale in quanto l’infezione è contratta precocemente ed è ampiamente diffusa nella popolazione. Tuttavia, la vaccinazione potrebbe essere efficace nel ridurre la frequenza e la gravità delle riattivazioni.
Varicella-zoster virus
Struttura
Alphaherpesviridae (Human Herpes virus 3)
Virione: morfologicamente simile a HSV (sferico) sebbene
dimensionalmente minore (d: 180 nm)
Peplos di origine cellulare, contiene la gran parte delle
glicoproteine viste in HSV:
▪ gB, gC, gH, gL: «concentrano» i virioni alla superficie della
cellula sensibile
▪ gC, gE, gI: proteggono i virioni dalla immunità umorale
(legando Complemento e Fc di Ig)
Tegumento (matrice): proteine virus-specifiche
Capsìde: icosaedrico, 100 nm, 162 capsomeri
Nucleoide: struttura a rocchetto con avvolto il DNA
Genoma: DNA ds (125 kbp), lineare; elevata omologia con HSVHerpesvirus: genoma The genomes of the herpesvirus are doubled-stranded DNA. The length and complexity of the genome differ for each virus. Inverted repeats in herpes simplex virus (HSV), varicella-zoster virus (VZV), and cytomegalovirus (CMV) allow the genome to recombine with itself to form isomers. Large genetic repeat sequences are boxed. The genomes of HSV and CMV have two sections, the unique long (UL ) and the unique short (US ), each of which is bracketed by two sets of inverted repeats of DNA. The inverted repeats facilitate the replication of the genome but also allow the UL and US regions to invert independently of each other to yield four different genomic configurations, or isomers. VZV has only one set of inverted repeats and can form two isomers. Epstein-Barr virus (EBV) exists in only one configuration, with several unique regions surrounded by direct repeats. Purple indicates direct repeat DNA sequences; green indicates inverted repeated DNA sequences. HHV6=human herpesvirus 6. [Medical Microbiology, 4th ed., Murray, Rosenthal, Kobayashi & Pfaller, Mosby Inc., 2002, Table 51-2]
Varicella-zoster virus Ciclo replicativo 1. VZV is presumed to use glycoproteins to bind cell surface receptors during attachment. 2. gB/gH-gL complex fuses virion and cell membranes either in endocytic vesicles or directly with plasma membrane. 3. Capsid is released into cytoplasm; at a nuclear pore, dsDNA genome is injected into the cell nucleus to initiate replication. 4. Glycoproteins are synthesized in the ER and trafficked to the Golgi during maturation, exocytosed then endocytosed and trafficked to the trans-Golgi network (TGN). 5. Some glycoproteins, including gB, are trafficked to the nuclear envelope (inner nuclear membrane, INM; outer nuclear membrane, ONM) and might have roles in egress from the nucleus. Glycoproteins are trafficked from the TGN for incorporation into nascent virus particles. (Oliver et al. Curr Clin Microbiol Rep. 2016 Dec; 3(4): 204–215)
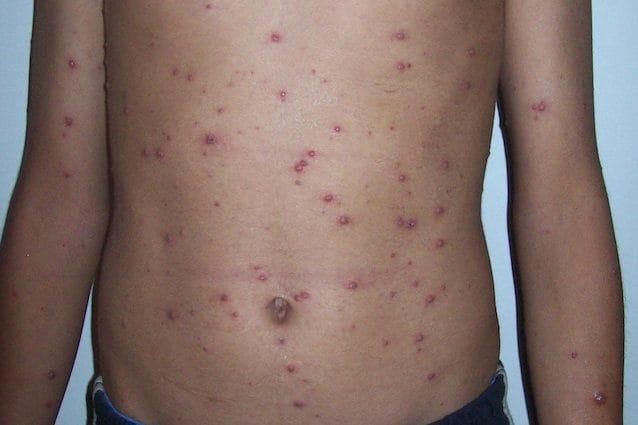
Varicella
Patogenesi – infezione primaria
1. Penetrazione nelle prime vie respiratorie o
congiuntiva mediata da aerosol e fluido di
vescicole
2. Replicazione nei linfonodi regionali
3. Viremia primaria (gg 4-6)
▪ febbre, malessere, cefalea
4. Replicazione in fegato, milza
5. Viremia secondaria (gg 11-13)
6. Diffusione a cute, organi di derivazione
ectodermica (sistema nervoso, alcune
ghiandole esocrine) e mucose
▪ lesioni cutanee secondarie a replicazione
endoteliale: rigonfiamento cellulare ed
accumulo di liquido interstiziale causano la
formazione di tipiche vescicole
Nell’ospite normoreattivo, l’infezione primaria
può essere asintomatica.Varicella
Clinica
▪ Fase iniziale: malessere generale e febbre.
▪ Comparsa di un eritema maculo-papulare, rosso e
pruriginoso, di solito prima sul ventre o sulla schiena e
viso, e poi si propaga a quasi tutto il corpo (cute e
mucose).
▪ Iniziale eruzione cutanea simile a brufoli o morsi di insetto,
quindi formazione di sottili vesciche piene di liquido chiaro,
che diventano torbide. La vescicola si rompe, lasciando
un’ulcerazione che si ricopre di una crosta.
▪ Le vesciche della varicella appaiono in più riprese da 2 a 4
giorni (presenza simultanea di lesioni a diverso stadio).
▪ Contagiosità del paziente: dalla comparsa del rash e fino
a 10-12 gg dopo.
▪ La guarigione è spontanea.
▪ Complicanze negli immunocompromessi:
▪ polmonite (principale serbatoio per la diffusione virale)
▪ encefalite erpetica (malattia neonatale)Varicella Patogenesi – infezione latente A seguito dell’infezione primaria e guarigione clinica il virus raggiunge le radici dorsali dei gangli sensoriali (cervicali, toracici e, a volte, lombo- sacrali) dove entra in uno stato di latenza che persiste per tutta la vita. Durante la fase di latenza, il virus può andare incontro a una serie di riattivazioni che, però, vengono contenute da una efficace risposta immunitaria (eventualmente richiamata da ulteriori contatti con il virus).
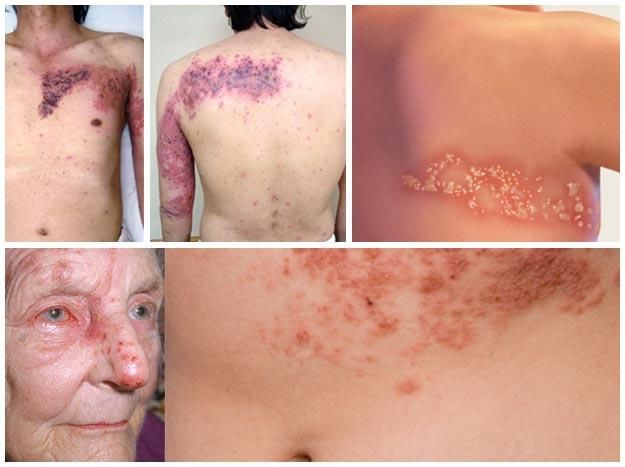
Zoster
Patogenesi – infezione secondaria (riattivazione)
Quando la risposta immunitaria dell’ospite è fortemente compromessa (trapiantati,
anziani, AIDS), è possibile la riattivazione del virus che torna a replicare e viene
rilasciato lungo tutta la via nervosa fino ad infettare la cute causando l’herpes zoster
(«fuoco di Sant’Antonio»). Tasso di riattivazione: 15-20% delle infezioni primarie.
L’infezione è di solito limitata all’innervazione cutanea di un singolo ganglio sensitivo
(dermatomero). Parestesia e bruciore precedono le lesioni cutanee di tipologia similare a
quelle della varicella.
La riattivazione avviene anche a distanza di anni ed è,
pertanto, più frequente nell’adulto.
Complicanze più comuni: neuralgia post-erpetica, encefalite,
zoster disseminato (potenzialmente fatale).
A volte la risposta immune riesce a controllare l’infezione a
livello cutaneo ma non a livello gangliare (infiammazione,
necrosi), risultando in «zoster sine herpete» (dolore senza Tra i gangli più colpiti il trigemino con la
sua branca oftalmica (zoster oftalmico)
lesioni cutanee).Varicella-zoster virus
Diagnosi di laboratorio
La diagnosi viene posta sulla base dell’evidenza clinica.
Isolamento del virus da colture cellulari esposte al liquido
delle vescicole (3-6 gg incubazione), con produzione di un
effetto citopatico simile a HSV (sincizi con formazione di
policariociti, corpi di Cowdry di tipo A).
Ricerca di antigeni virali, mediante anticorpi fluorescenti, in VZV: corpi di Cowdry di tipo A
materiale da raschiamento delle lesioni cutanee.
Evidenziazione di DNA virale mediante PCR in liquido da
vescicole, biopsie, croste.
Titolazione anticorpale mediante immunoenzimatica (ELISA)
o immunofluorescenza diretta.
VZV: immunofluorescenzaVaricella-zoster virus
Terapia e profilassi
TERAPIA
▪ Terapia soltanto negli immunocompromessi. Nei bambini il
decorso è benigno.
valaciclovir
▪ Farmaco di elezione per varicella: aciclovir (analogo aciclico della
guanosina).
▪ Farmaco di elezione per zoster è valaciclovir (profarmaco di
aciclovir; convertito nel fegato), quindi aciclovir e famciclovir
(analogo aciclico della adenina). famciclovir
▪ Brivudin, un analogo nucleosidico pirimidinico, ha buona attività
nelle polmoniti da varicella.
brivudin
PROFILASSI
▪ Immunizzazione passiva: somministrazione di Ig anti-VZV ha
effetto protettivo.
▪ Immunizzazione attiva (vaccino): virus attenuato da somministrare
dopo i 2 anni di età.Herpesviridae (Betaherpesvirinae)
Proprietà caratteristiche degli herpesvirus umani
From Medical Microbiology, 4th ed., Murray, Rosenthal, Kobayashi & Pfaller, Mosby Inc., 2002, Table 51-1.
Alphaherpesvirinae: ampio spettro d’ospite (inoculazione per scarificazione o per via
intracerebrale), si replicano in diversi tipi di colture cellulari con caratteristico effetto citopatico e
formazione di inclusioni nucleari.
Gammaherpesvirinae: specie-specifici, si replicano solo in colture cellulari della specie sensibile.
Betaherpesvirinae: ristretto spettro d’ospite, tropismo per le cellule linfoidi.Beta-Herpesvirinae ▪ Spettro d’ospite più ristretto ▪ Ciclo riproduttivo più lento ▪ In vitro l’infezione progredisce lentamente e le cellule assumono un aspetto rigonfio (citomegalia). ▪ Nell’ospite naturale infettano le cellule linfoidi, e sono latenti nelle ghiandole secretorie, nelle cellule linfo-reticolari, nel fegato e altri tessuti. ▪ Non si osserva neurotropismo. ▪ Nell’uomo: Citomegalovirus umano (HCMV), Herpesvirus umano tipo 6 (HHV-6), Herpesvirus umano tipo 7 (HHV-7). ▪ Negli animali: Citomegalovirus murino (MCMV).
Citomegalovirus
Struttura
Virione: sferoidale, d: 250 nm (il più grande tra herpesvirus)
Peplos di origine cellulare, contiene glicoproteine (g):
▪ gB, gH: ruolo preminente per adsorbimento (a
proteoglicani di superficie cellulare) e penetrazione
previa interazione con il recettore per EGF (Epidermal
Growth Factor)
▪ recettori cellulari: EGFR, integrine, PDGFRα,
BST/tetherina
Tegumento (matrice): proteine virus-specifiche regolatrici la
espressione a cascata dei geni , , γ
Capsìde: icosaedrico, 100 nm, 162 capsomeri
Genoma: DNA ds (240 kbp), lineare
Cytomegalovirus progeny into an infected cell, TEMHerpesvirus: genoma The genomes of the herpesvirus are doubled-stranded DNA. The length and complexity of the genome differ for each virus. Inverted repeats in herpes simplex virus (HSV), varicella-zoster virus (VZV), and cytomegalovirus (CMV) allow the genome to recombine with itself to form isomers. Large genetic repeat sequences are boxed. The genomes of HSV and CMV have two sections, the unique long (UL ) and the unique short (US ), each of which is bracketed by two sets of inverted repeats of DNA. The inverted repeats facilitate the replication of the genome but also allow the UL and US regions to invert independently of each other to yield four different genomic configurations, or isomers. VZV has only one set of inverted repeats and can form two isomers. Epstein-Barr virus (EBV) exists in only one configuration, with several unique regions surrounded by direct repeats. Purple indicates direct repeat DNA sequences; green indicates inverted repeated DNA sequences. HHV6=human herpesvirus 6. [Medical Microbiology, 4th ed., Murray, Rosenthal, Kobayashi & Pfaller, Mosby Inc., 2002, Table 51-2]
Citomegalovirus: ciclo replicativo (A) Virions enter the cell: gB recognizes cell proteoglicans for the viral concentration at the cell surface, and then gH mediates cell penetration linking to the EGF-receptor. Two ways of entry. Tegument and capsid proteins are delivered to the cytoplasm. (B) The capsid travels to the nucleus, then the genome is delivered and circularized. Tegument proteins regulate host cell responses and initiate the temporal cascade of the expression of viral immediate early (IE) genes, followed by delayed early (DE) genes, which initiate viral genome replication, and late (L) genes. (C) Late gene expression initiates capsid assembly in the nucleus, followed by nuclear egress to the cytosol. Capsids associate with tegument proteins in the cytoplasm and are trafficked to the viral assembly complex (AC) containing components of the endoplasmic reticulum (ER), Golgi apparatus, and endosomal machinery. The capsids further acquire tegument and viral envelope by budding into intracellular vesicles at the AC. (D) Enveloped infectious particles are released along with non-infectious dense bodies. (Adapted from Jean Beltran & Cristea, 2014)
Citomegalovirus (CMV)
Replicazione ed effetto citopatico
La replicazione ha durata maggiore (8-14 giorni) vs altri
herpesvirus.
In vitro si replica soltanto in fibroblasti umani, sebbene in
vivo possa essere isolato anche da altri tipi cellulari (pancreas,
endotelio, ghiandole salivari, rene).
L’infezione causa un caratteristico aumento volumetrico della
cellula (citomegalìa) con formazione di inclusioni intranucleari
(classiche degli herpesvirus) e caratteristiche inclusioni
citoplasmatiche ad «occhi di civetta» a significato
patognomonico.
Causa lesioni focali (come in VZV) perché non viene rilasciato
nel mezzo colturale ma diffonde direttamente a cellule contigue.
La cellula va incontro a morte per lisi.Citomegalovirus (CMV)
Meccanismi patogenetici
Infezione primaria → infezione persistente latente per tutta
la vita dell’individuo infetto.
L’infiammazione può portare a riattivazione di CMV nei
monociti che, sotto l’azione delle citochine proinfiammatorie,
si attivano a macrofagi. Il virus può così diffondere ad
altre cellule.
Ruolo potenziale in: malattie cardiovascolari, autoimmuni e
alcune neoplasie.
Patologie vascolari: Attivazione monociti → riattivazione di
CMV. Adesione macrofagica alle cellule endoteliali: CMV
influenza il metabolismo lipidico, portando allo sviluppo di
cellule schiumose. La sua trasmissione all’endotelio
contribuisce alla crescita della placca, aumenta
l’infiammazione e contribuisce alla comparsa di sintomi
ischemici (aumentata espressione di molecole di adesione
ed aumento di transmigrazione leucocitaria e
dell’infiammazione), aumenta il rilascio del fattore di von
Willebrand (vWF) portando all’attivazione delle piastrine e
alla loro aggregazione: trombi e ischemie.Citomegalovirus (CMV)
Possibili esiti dell’infezione
▪ La possibile malattia dipende
pesantemente dallo stato immunologico
del paziente
▪ Ampio tropismo dovuto al
riconoscimento, da parte di gB e gC, di
eparan-solfato (molecola ubiquitaria)
come recettoreCitomegalovirus (CMV)
Manifestazioni cliniche
Patogeno opportunista. Infezione molto diffusa
(sieroprevalenza 70%) e generalmente
asintomatica.
Trasmissione dell’infezione per contatto diretto
con diversi materiali biologici (sangue, saliva,
sperma); orizzontale e verticale (sia per infezione
primaria che per riattivazioni).
Periodo incubazione (4-8 settimane).
Infezione primaria (acquisita nella prima infanzia)
è asintomatica nel normoreattivo. Liberazione
costante di virus per anni con salive e urine.
In pochi casi, invece, si manifesta come una
sindrome simil-mononucleosica (malessere
generalizzato, febbre, faringite, linfoadenopatia)
con coinvolgimento epatico.
Virus presente in saliva, sangue, urine, sperma,
secrezioni lacrimali.Citomegalovirus (CMV)
Manifestazioni cliniche
▪ Infezione connatale (in gravidanza): prima
infezione (30-40%), riattivazione (II-III semestre;
0.5-2%), re-infezione. Trasmissione per via
transplacentare dell’infezione al feto (1% neonati
vivi); possibile (10%) malattia citomegalica con
inclusioni (porpora trombocitopenica grave,
miocardite, polmonite, sordità, danni irreversibili al
SNC, microcefalia).
Mortalità (30%) o gravi reliquati nei sopravvissuti
(epatosplenomegalia, microcefalia e calcificazioni,
sordità, ritardo mentale).
▪ Contagio all’atto del parto (infezione perinatale)
per contatto con secrezioni vaginali o latte materno
(durante l’allattamento).
Quadro clinico meno grave (ipoacusia, polmonite)
per trasferimento passivo di anticorpi materni.
▪ L’infezione può essere contratta anche a seguito di
trasfusione con sangue da donatori con infezione
silente. Sindrome post-perfusionale (interventi
chirurgici con circolazione extracorporea).Citomegalovirus (CMV)
Manifestazioni cliniche
A seguito di infezione primaria, CMV si localizza a
livello di endotelio, nei leucociti del sangue
periferico, nel midollo dove rimane in fase
quiescente all’interno di cellule mononucleate,
che rappresentano un serbatoio per le riattivazioni.
In presenza di immunocompromissione, trapianto
d’organo, uso di corticosteroidi e/o
immunosoppressori, il quadro clinico è causato da
una riattivazione di un’infezione primaria.
Polmonite interstiziale con febbre elevata
(complicanza più grave e frequente).
Fenomeni di rigetto nei trapiantati.
Riattivazione silente in donna gravida (II e III
trimestre gestazione). esofagite colite
Nei malati di AIDS, infezione disseminata
(corioretinite, esofagite, gastroenterite, colite).
corioretiniteCitomegalovirus (CMV)
Diagnosi
Isolamento virale in colture di fibroblasti umani a partire da
saliva, lavaggio bronco-alveolare, urine, leucociti. Effetto
citopatico caratteristico ad «occhio di gufo», ma richiede almeno
10-14 giorni (lenta replicazione).
CMV: CPE in fibroblasti
Ricerca rapida di antigeni virali mediante IF:
- pp65 (di tegumento) nei leucociti (antigenemia)*
- antigeni precoci (p72/p86) nei fibroblasti
IF assay: CMV pp65 in leucociti
Ricerca rapida di sequenze geniche:
- n copie DNA virale mediante PCR quantitativa (viral load)*
- ibridazione in situ con sonde a DNA (biopsie)
* utile per il monitoraggio della possibile riattivazione dell’infezione in In situ DNA-hybridization: tessuto fissato
immunodepressi (AIDS, trapiantati) in formalina ed incluso in paraffinaCitomegalovirus (CMV) Diagnosi – infezione connatale e perinatale Screening sierologico: ricerca anticorpale mediante reazione di neutralizzazione o fissazione del Complemento. Infezione connatale: ▪ IgG+, infezione pregressa ▪ IgM+, infezione primaria acuta o recente (in gravidanza e immunocompromessi) Causa possibili falsi-positivi, il riscontro di IgM deve essere confermato con altri test (es. test di avidità IgG, amniocentesi, analisi sangue fetale). Infezione perinatale: ricerca di virioni nel neonato (primi 21 gg vita) in urine, saliva, sangue
Citomegalovirus (CMV)
Terapia
ganciclovir foscarnet
Ganciclovir e Foscarnet per le infezioni gravi in pazienti immunocomopromessi.
Meccanismo azione:
▪ Ganciclovir: nucleoside analogo della 2-deossiguanosina; agisce da
profarmaco: deve essere fosforilato (da fosfotransferasi virale) per inibire la DNA-
polimerasi e causare la terminazione della catena di DNA.
▪ Foscarnet: analogo organico del pirofosfato inorganico, esercita la sua attività
antivirale tramite una inibizione selettiva sul sito di legame del pirofosfato
della DNA polimerasi virus-specifica già a concentrazioni che non influenzano la
DNA polimerasi della cellula ospite. Utilizzato in caso di resistenza a ganciclovir,
sebbene siano stati osservati ceppi resistenti anche a foscarnet (a seguito di
alterazioni a carico della DNA polimerasi).
Non esistono trattamenti prenatali efficaci e sicuri per prevenire la trasmissione
madre-feto dell’infezione né per ridurre le conseguenze di un’infezione congenita.
Ganciclovir e foscarnet sono estremamente dannosi per il feto.Citomegalovirus (CMV)
Controllo
▪ Non esiste attualmente un vaccino per la prevenzione del CMV.
▪ Il modo migliore per limitare il rischio di contagio è un’attenta igiene personale,
soprattutto per le categorie di persone più vulnerabili alla malattia (donne in
gravidanza, individui immunodepressi, bambini piccoli o appena nati):
▪ lavare le mani prima di mangiare, preparare e servire il cibo, dopo aver
cambiato i bambini, dopo essere andati in bagno e dopo ogni tipo di contatto
con fluidi corporei
▪ evitare di scambiarsi posate o altri utensili durante i pasti, soprattutto con
bambini piccoli
▪ pulizia della casa, soprattutto delle superfici contaminate da fluidi corporei
(come saliva, urina, feci, liquidi seminali e sangue)Questo materiale non può essere distribuito, modificato o pubblicato né in forma cartacea, né su un sito, né utilizzato per motivi pubblici o commerciali. E’ possibile utilizzare il materiale solo per motivi personali e non commerciali, purché ogni copia di questo materiale preservi tutti i diritti di copyright e di proprietà intellettuale, sempre a seguito di formale richiesta rivolta all’Autore (Prof. Giovanni Di Bonaventura; gdibonaventura@unich.it)
Puoi anche leggere

















































