Studio dei polimorfismi del DNA in Vitis Vinifera per la discriminazione di cloni
←
→
Trascrizione del contenuto della pagina
Se il tuo browser non visualizza correttamente la pagina, ti preghiamo di leggere il contenuto della pagina quaggiù
Final Report
“Studio dei polimorfismi del DNA in Vitis Vinifera per la
discriminazione di cloni”
Coordinatore Scientifico: Dott. Donato Antonacci (CRA-UTV Turi)
Borsista: Dott. Angelo Anaclerio
RIASSUNTO
L’Aglianico è un antico vitigno, diffuso soprattutto nel Mezzogiorno d’Italia e coltivato in
modo particolare in Campania, Basilicata ed in misura inferiore in Puglia e Molise (Antonacci D.,
2004). E’ caratterizzato da un’elevatissima variabilità intravarietale e la sua importanza è
testimoniata dal fatto che rientra per la produzione di vini in alcune IGT, DOC e DOCG. Lo scopo
del presente lavoro è stato quello di verificare, attraverso l’utilizzo della metodica molecolare AFLP,
la possibilità di discriminare cloni o accessioni appartenenti alla cultivar Aglianico. Per tale
obiettivo abbiamo considerato complessivamente 31 biotipi tra Aglianico n. (cod. 002) e Aglianico
del Vulture n.(cod. 266), iscritti nel Registro Nazionale delle Varietà di Vite (D.P.R. 24 dicembre
1969, n° 1164) come due diverse cultivar, ma che in realtà come dimostrato in letteratura con i
marcatori SSR sembrano essere la medesima varietà. Due cloni certificati, uno per l’Aglianico n. e
uno per l’Aglianico del Vulture n. sono stati utilizzati come confronto e riferimento e pertanto
inseriti nell’analisi. L’utilizzo di 30 descrittori morfologici, 13 marcatori SSR e 10 combinazioni
AFLP ha permesso di investigare la diversità esistente all’interno della popolazione di Aglianico
raccolta nel Sud Italia. Differenze significative per i descrittori morfologici sono state rilevate
esclusivamente a livello della foglia matura, mentre poche o alcuna a livello della bacca, grappolo e
germoglio. Una completa uniformità genetica è stata rilevata con i marcatori SSR per tutti i 13 locus.
Anche gli AFLP hanno evidenziato un elevato grado di similarità tra le accessioni e tra i due cloni
certificati, tuttavia a differenza delle altre metodiche di caratterizzazione hanno permesso di
distinguere ogni singolo individuo esaminato. Appare evidente quindi il maggiore potere
discriminatorio della metodica AFLP nella discriminazione di individui all’interno di una varietà.
Tuttavia questa analisi al momento non può prescindere dalla corretta attribuzione varietale
mediante SSR e da una previa analisi fenotipica dei campioni da esaminare. Tuttavia non è da
escludere che ulteriori analisi effettuate con una metodica AFLP standardizzata possano portare ad
una simultanea identificazione di varietà e discriminazione di cloni nella stessa indagine.
INTRODUZIONE
L’Aglianico è un vitigno diffuso soprattutto nel Mezzogiorno d’Italia, coltivato in modo
particolare in Campania (circa 6700 ha), Basilicata (circa 2600 ha) ed in misura inferiore in Puglia e
Molise (ISTAT, 2000). Nel panorama ampelografico, senza alcun dubbio, il vitigno Aglianico è da
annoverare tra le varietà più rappresentative e storiche dell’Italia meridionale. Infatti, non è solo uno
dei più diffusi vitigni nelle aree a propensione viticola, ma è anche quello più ricco di citazioni e di
riferimenti storici.
L’origine dell’Aglianico è controversa; numerose sono state le tesi avanzate dagli studiosi di
letteratura enoica, alcune di queste sono recentissime. A lungo si è ritenuto che l’Aglianico si
identificasse con le antiche uve Helleniche. Tale tesi è stata sostenuta per primo da Columella
Onorati (1806) e ripresa da Carlucci (1909), che ha studiato a fondo l’Aglianico e realizzato la
prima monografia completa sul vitigno. Carlucci, in particolare, sostiene che l’Aglianico si
identifichi con il vitigno che, noto con il nome di Ellenica o Ellanica, veniva utilizzato, in epoca
1romana, per produrre il Gaurano, vino derivante dall’area del Massico, che oggi coincide con il
Monte Barbaro, il Faustiano e con i colli Falciano. Inoltre Carlucci nel suo lavoro fa derivare il
termine Aglianico da una corruzione di Hellenico, prima in Hellenica e, successivamente, in epoca
aragonese (XV sec.), in Aglianico, mediante la trasformazione della doppia elle nel suono “gl”.
Questa tesi è stata suffragata anche nelle opere di Giovan Battista Porta (1581) e di Granata (1883).
Nonostante numerose ed autorevoli adesioni a tale tesi avanzata da Carlucci, negli ultimi anni sono
state elaborate ipotesi diverse. Murolo (1970) avanza l’ipotesi che Aglianico possa derivare dai
termini greci “aglaos” (chiaro) e “aglaio” (splendore): e cioè Aglianico come vino rubino e
splendente (Vitagliano, 1989).‟ultimaL congettura è recen tissima ed è stata avanzata da
Piancastelli (2003). Le radici del termine Aglianico sarebbero da ricercarsi nella giovanile asprezza
e astringenza del vino, per cui deriverebbero da “aglucos”, cioè senza zucchero, cioè amaro. Tale
tesi affascina Moio (2003) in quanto è confortata da alcune caratteristiche peculiari dell’uva
Aglianico, che presenta una elevata acidità tollerabile, con conseguente maggiore reattività delle
forme polifenoliche monomere che si traducono in una più accentuata percezione dell’attributo
amaro.
Le numerose opinioni, pur essendo diverse, concordano tutte nel riconoscere all’Aglianico
una nobile storia e una millenaria presenza soprattutto in Campania e più in generale nell’Italia
meridionale, non è un caso quindi, che dalle uve di Aglianico si ottengano i vini campani più
affermati e prestigiosi e che rientri in un gran numero di vini IGT e DOC.
La stessa storia millenaria dell’Aglianico ha portato nel tempo alla formazione di diversi
biotipi che hanno dato origine ad un numero elevatissimo di sinonimi con cui il vitigno è indicato
nelle diverse aree viticole ove è presente. Tra i sinonimi ricordiamo: Aglianica, Aglianica vera,
Aglianica Cola Giovanni, Glianica, Glianico, Gnanica, Ellanico, Aglianicone, glianica bastarda,
Aglianichella di S. Severino, Ellenico, Aglianico zerpoluso, Aglianico mascolino, Cascavoglia,
ecc….
Per l’Aglianico così come per tutte la varietà di vite, evidenze e citazioni storiche, integrate
da dati morfologici (ampelografia), sono state il primo strumento per caratterizzare le varietà e
presumere parentele. Tuttavia il crescente verificarsi di contestazioni nell’attribuzione varietale
hanno spostato l’attenzione a livello genetico (Labra et al., 2004). I marcatori molecolari
maggiormente utilizzati in ambito viticolo sono attualmente gli SSR (simple sequence repeats),
efficacemente utilizzati per l’identificazione di cultivar (Bowers et al. 1999; Crespan 2003; This et
al. 2004; Jahnke et al. 2009; Zoghlami et al. 2009) e per la discriminazione clonale (Regner et al.
2000; Riaz et al. 2002; Bertsch et al. 2003; Crespan 2003; Hocquigny et al. 2004; Gonzàles-Techera
et al. 2004). Tuttavia numerose sono anche le ricerche in cui viene evidenziata l’impossibilità di
distinguere cloni di vite attraverso l’utilizzo degli SSR (Crespan and Milani 2001; Imazio et al.
2002; Vignani et al. 2002; Ibànez et al. 2003). Imazio et al. (2002) consigliano la metodica AFLP
(amplified fragments length polymorphism) rispetto agli SSR per discriminare cloni appartenenti
alla medesima varietà o per ricercare variabilità tra varietà affini. Risultati positivi con la metodica
AFLP sono stati riportati da (Vignani et al. 2002; Labra et al. 2004; Blaich et al. 2007; Stenkamp et
al. 2009).
MATERIALI E METODI
Materiale vegetale
Nell’ambito del progetto Vitivin-Valut, inerente anche il recupero e la valorizzazione dei
vitigni autoctoni, sono state recuperate 110 accessioni di Aglianico (74 accessioni di Aglianico n. e
36 di Aglianico del Vulture n). Lo scopo inizialmente è stato quello di recuperare il maggior numero
di biotipi riscontrabili sui territori esplorati in modo da salvaguardarne la biodiversità.
2Successivamente queste accessioni, opportunamente innestate, sono state caratterizzate con
descrittori morfologici e marcatori molecolari con il fine di verificare se appartenessero alla
cultivar Aglianico.
Per lo svolgimento del lavoro di borsa, su tutte queste accessioni è stata effettuata una prima analisi
fenotipica, visiva, al fine di individuare quei campioni o biotipi che presentavano caratteristiche
distintive rispetto agli individui della collezione. Sono stati così scelti 31 biotipi tra Aglianico n. e
Aglianico del Vulture. Due cloni certificati Aglianico n. (IRGV Codice 002) e Aglianico del Vulture
n. (IRGV Codice 266), presenti nello stesso campo sperimentale sono stati utilizzati come
riferimento. Complessivamente 33 sono i genotipi considerati.
I campioni esaminati sono localizzati nel campo sperimentale del CRA-UTV di Turi e ognuno di
essi è stato replicato 10 volte. La distanza tra i filari è di 2,5 metri, mentre quella tra le piante è di 1
m.
Analisi Ampelografiche
Tra il 2007, 2008 e 2009 sulle 31 accessioni e sui 2 cloni certificati sono stati misurati 30
caratteri morfologici, seguendo la lista dei descrittori sviluppata dall’Organizzazione Internazionale
della Vigna e del Vino (OIV, 2ND Edition 2007). Tra i caratteri sono stati inclusi anche quelli
minimi per uno studio preliminare su radici, foglie, grappoli e acini (Tab. 1). Le misure su grappoli
e acini sono state effettuate considerando rispettivamente la media di 20 grappoli e di 50 acini.
Analisi Genotipiche
Il DNA genomico totale è stato estratto da foglie giovani (nella fase di fioritura della pianta)
utilizzando un kit commerciale e seguendo il protocollo consigliato dalla casa produttrice (Dneasy
Plant; Qiagen, Hilden, Germany). La qualità e la quantità del DNA sono state misurate attraverso
corsa elettroforetica su gel di agarosio allo 0,8% e analisi spettrofotometrica a 260 nm.
Sono stati analizzati 13 loci microsatellite, sei dei quali richiesti dal EU project Genres CT96 N° 81:
VVS2, VVMD5, VVMD7, VVMD27, VrZAG62 e VrZAG79 (This et al., 2004); mentre i restanti
sette sono stati scelti tra i più polimorfici sulla base dei dati di letteratura: VVMD28, VVMD25 e
VVMD32 (Bowers et al., 1999) ISV2, ISV3, ISV4 e VMCNG4b9 (Crespan 2003). La Multiplex
PCR (Polimerase Chain Reaction) è stata condotta utilizzando l’Eppendorf Mastercycler® EP
thermocycler (Eppendorf, North America, Inc., New York, NY). Per ogni campione è stata allestita
una Mix costituita da: Mix Qiagen (contenente Dna polimerasi stabilizzata, dNTP, Mg2+), primer e
H2O per raggiungere il volume finale di 20 µl (Qiagen, Hilden, Germany). Due o tre coppie per
volta sono state combinate per co-amplificare in una singola reazione. Il primer “forward” è stato
marcato con WellRED dyes, D2-PA (black), D3-PA (green) or D4-PA (blue), all’estremità 5’finale.
I cicli di amplificazione prevedevano: un iniziale step di attivazione delle polimerasi a 95°C per 5
min, seguivano 35 cicli con denaturazione a 98°C per 5 s, annealing a 55°C per 30 s, estensione a
68°C per 9 s, e una estensione finale a 72 C per 1 min. I prodotti di PCR sono stati analizzati con il
sequenziatore CEQ™ 8000 Series Genetic Analysis System, utilizzando uno standard di peso
molecolare CEQ DNA Standard Kit Size 400 (Beckman Coulter S.p.A., Milan, Italy). Le
dimensioni dei frammenti sono state stimate attraverso analisi automatizzata.
L’analisi AFLP è stata condotta seguendo il protocollo riportato da Vos et al. (1995) con alcune
modifiche. Il DNA genomico (150 ng) ha subito una doppia digestione per 1 h con 20 unità di
EcoRI and 10 unità di MseI. I frammenti di DNA sono stati ligati con 3.5 unità di T4 ligasi a un
3adattatore EcoRI (5 pmol) e ad un adattatore MseI (50 pmol) in un volume finale di 50 μl a 15°C
overnight.
Venticinque µl derivanti dalla digestione e ligazione sono stati usati come templato in una reazione
di pre-amplificazione con l’aggiunta di 10x PCR buffer Fermentas, 25 mM MgCl2, 250 ng di
primer EcoRI (+1N) e 50 ng di primer MseI (+1N), 10 mM di dNTP, 1.5 unità di Taq DNA
polymerase, in un volume finale di 45 μl. L’amplificazione ha previsto: 3 min a 94ºC, 21 cicli di 30
s a 94ºC, 1 min a 56ºC, e 1 min a 72ºC, con uno step finale di estensione a 72ºC per 7 min. I
prodotti della pre-amplificazione sono stati valutati su gel di agarosio all’1%. Successivamente, i
campioni sono stati diluiti 1:10 o 1:20 in base alla loro iniziale concentrazione. Quattro μl dei
prodotti di pre-amplificazione sono stati utilizzati come templato per l’amplificazione selettiva.
Sono stati aggiunti 10x PCR buffer Fermentas, 25 mM MgCl2, 16 ng primer EcoRI e 96 ng primer
MseI, 10 mM dNTP, 0.4 unità Taq DNA polymerase, in un volume finale di 10 μl. I primer EcoRI
sono stati legati con fluorocromi FAM o HEX. Le condizioni di PCR utilizzate sono state le
seguenti: 94ºC per 3 min, 12 cicli di 30 s a 94ºC, 30 s a 65ºC (la temperature annealing è stata
ridotta ad ogni ciclo di 0.7ºC) e 1 min a 72ºC; 23 cicli addizionali completano la fase di
amplificazione selettiva: 30 s a 94ºC, 30 s a 56ºC e 1 min at 72ºC. Le sequenze di adattatori, primer
per l’amplicazione pre-selettiva e selettiva sono riportati nella Tabella 2. Un totale di 10
combinazioni di primer con 3 nucleotidi selettivi sono state utilizzate. I prodotti di amplificazione
sono stati prima denaturati a 94°C per 5 min e successivamente separati mediante elettroforesi
capillare utilizzando il sequenziatore ABI PRISM ® 3100 Avant Genetic Analyzer (Applied
Biosystems) con GS ROX 500 come standard interno. Gli elettroferogrammi AFLP sono stati
acquisiti e analizzati con il Software Genographer 2.1.4
(http://sourceforge.net/projects/genographer), che permette di ricostruire una virtuale corsa
elettroforetica su gel, con bande suddivise in base al proprio peso molecolare. Le letture delle
diverse combinazioni di primer sono state effettuate indipendentemente da due operatori, e da
queste sono state considerate solo quelle bande non ambigue.
RISULTATI
La comparazione dei dati ampelografici per i descrittori primari tra i cloni certificati
Aglianico n. e Aglianico del Vulture n. ha mostrato differenze morfologiche solo relative alle foglie
mature, mentre completa uniformità è stata riscontrata per i descrittori dei giovani germogli e dei
grappoli (Tab. 1). In particolare a livello della foglia adulta l’Aglianico del Vulture n. differisce
dall’Aglianico n. per i seguenti descrittori OIV:
cod.067 (carattere forma del lembo): orbicolare vs. pentagonale;
cod.068 (carattere numero lobi): tre vs. cinque;
cod.076 (carattere forma dei denti): su entrambi i lati convessi vs. su entrambi i lati rettilinei;
cod.084-087 (carattere peli eretti): l’Aglianico del Vulture n. mostra una maggiore presenza peli
eretti sulle nervature principali della pagina inferiore del lembo rispetto all’Aglianico n..
Le 31 accessioni analizzate per i descrittori primari hanno mostrato maggiore variabilità rispetto ai
cloni certificati. In particolare oltre ad evidenziare differenze a livello della foglia adulta è stata
rilevata variabilità anche per il descrittore del germoglio (cod. 004) che ha evidenziato, per 4
accessioni, una densità dei peli striscianti inferiore rispetto ai cloni di riferimento e paragonabile a
quella della Garnacha tinta N..
4Sei descrittori primari hanno mostrato completa uniformità sia tra i cloni certificati che tra le
diverse accessioni: completa apertura dell’apice (cod. OIV 001), 2 o nessun viticcio consecutivo
(cod. OIV 016), colore verde della pagina superiore del lembo della quarta foglia (cod. OIV 051),
presenza di denti del seno peziolare su entrambi i lati delle foglie mature (cod. OIV 81-2), grappoli
di forma globosa (cod. OIV 223) e colore della buccia (cod. OIV 225) blu-nera.
Poiché maggiormente influenzati dall’ambiente, i descrittori secondari (Tab. 2), hanno
mostrato una più ampia variabilità tra i cloni certificati e tra le accessioni esaminate. Questa
biodiversità ha riguardato non solo i descrittori della foglia matura ma anche i descrittori del
grappolo e del germoglio, mentre i descrittori delle bacche non hanno mostrato variabilità. In
Aglianico del Vulture n. i grappoli in media, rispetto all’Aglianico n., hanno evidenziato maggiore
lunghezza (cod. OIV 202), larghezza (cod. OIV 203), media densità (cod. OIV 204), con un più
lungo peduncolo (cod. OIV 206) e una forma conica vs. forma cilindrica (cod. OIV 208). I grappoli
variavano dagli 8.0 cm ai 20.0 cm di lunghezza (OIV 202) con larghezza compresa tra i 4.0 e 16.0
cm (OIV 203). La compattezza variava da media a spargola (OIV 204), mentre la lunghezza del
penducolo del grappolo primario varia dai 0.3 cm ai 0.9 cm (OIV 206). Tutti i grappoli avevano
forma cilindro-conica (OIV 208).
Inoltre l’Aglianico del Vulture n. ha manifestato a livello della foglia, rispetto all’Aglianico n., una
maggiore dimensione del lembo (cod. OIV 067), denti più corti (cod. OIV 078), forma della base
del seno peziolare a “V” rispetto a quella dell’Aglianico ad “U”.
I caratteri presenza di denti del seno peziolare (cod. OIV 081-1) e presenza di denti nei seni laterali
superori rappresentano un ulteriore elemento di discriminazione tra le accessioni ma non tra i cloni
certificati. La sola accessione afferente al biotipo Sannio ha mostrato una maggiore intensità della
pigmentazione antocianica dei peli striscianti dell’apice (cod. OIV 003).
Quattro descrittori secondari (cod. OIV 209, 231, 235 e 241) hanno mostrato valori paragonabili tra
tutti gli individui esaminati. In particolare i descrittori OIV cod. 231, 235 e 241 sono descrittori
dell’acino.
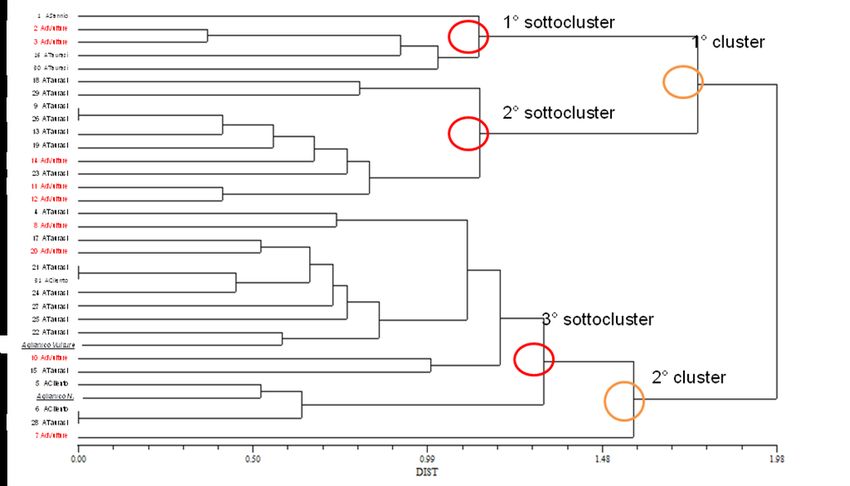
I dati ampelografici sono stati utilizzati per ottenere un dendrogramma di similarità (Fig. 1)
attraverso il software NTSYS, utilizzando il coefficiente di distribuzione Dist e l’algoritmo
UPGMA. Il dendrogramma mostra la formazione di 2 cluster principali che originano 4 sotto-
cluster. Nel terzo sotto-cluster si ritrovano le cultivar certificate, mentre il quarto sotto-cluster è
costituito dalla sola accessione 7, biotipo Vulture. Appare evidente l’impossibilità di discriminare
tutte le accessioni, in quanto le accessioni 9 e 26, 21 e 31 ed infine 6 e 28, nonostante i 30 caratteri
fenotipici esaminati mostrano uno stesso fenotipo. Inoltre dall’analisi si evince un mancato
raggruppamento tra le accessioni afferenti alle cultivar certificate rafforzando così la tesi secondo la
quale l’Aglianico n. e l’Aglianico del Vulture n. sarebbero in realtà la stessa varietà.
Nonostante i dati ampelografici siano stati prelevati sempre dagli stessi operatori e sulle
stesse piante poste nelle medesime condizioni di allevamento, non si possono escludere errori di
rilevazione del carattere e una certa pressione ambientale alla quale le diverse accessioni sono
sottoposte. Per ovviare a questo inconveniente e confermare l’origine genetica delle differenze
morfologiche riscontrate sono stati esaminati alcuni polimorfismi del DNA che notoriamente non
sono influenzabili dall’ambiente.
L’analisi SSR ha rivelato una totale uniformità, stesso profilo per ogni singolo locus, tra
tutte le accessioni e le cultivar certificate per i 13 loci testati (Tab. 3). Due (VrZAG62, VVMD7)
dei sei (VrZAG79, VVS2, VVMD27 e VVMD5) loci richiesti dall’EU project Genres CT96 N° 81
5“European network for grapevine genetic resources conservation and characterisation”, sono
risultati monomorfici (omozigosi), mentre gli altri quattro loci hanno mostrato un profilo
eterozigote. Anche gli altri sette marcatori (VVMD28, VVMD25, VVMD32, ISV2, ISV3, ISV4 e
VMCNG4b9) scelti poiché tra i più polimorfici facendo riferimento ai dati di letteratura, hanno
mostrato un profilo eterozigote. Tramite l’utilizzo dei 13 marcatori SSR è dunque impossibile
distinguere l’Aglianico n. e l’Aglianico del Vulture n. ed è pertanto ipotizzabile che le due cultivar
siano in realtà la medesima varietà nonostante le differenze fenotipiche delle cultivar certificate.
Le 10 combinazioni di primer AFLP (Tab. 4) testate sulle 31 accessioni e sui 2 cloni
certificati hanno prodotto complessivamente 377 bande, delle quali 108 sono risultate polimorfiche.
Il polimorfismo è stato inteso come presenza o assenza di una determinata banda nella ricostruzione
virtuale del gel da parte del Software Genographer 2.1.4. Questo Software ha permesso di
standardizzare le letture, attraverso la possibilità di selezionare l’intensità del segnale e di scegliere
l’intervallo di lettura del polimorfismo; ed inoltre ha permesso di evitare tutti quegli errori e rischi
derivanti dalla lettura dei gel in cui i primer sono marcati con radioattivo. Ulteriore vantaggio
fornito dal Software è stato quello di aver poter effettuare le letture nel tempo.
Sono state lette solo quelle bande di indubbia lettura e con un peso molecolare non inferiore alle
100 bp. In figura 2 è riportato l’ esempio di una combinazione di primer (E44-M59) ottenuta con il
Software Genographer. Il numero di bande ottenute per singola combinazione è stato variabile e
comunque compreso tra un minimo di 25 e un massimo di 61; mentre il numero di bande
polimorfiche è risultato compreso tra 4 e 19. Dalla lettura è stata originata una matrice di similarità,
che attraverso NTSYS, utilizzando il coefficiente di similarità DICE e l’algoritmo UPGMA, ha
generato un dendrogramma di similarità per i dati AFLP (Fig. 3). Appare evidente l’elevato livello
di similarità tra le accessioni, superiore al 94%, e per i due cloni certificati che hanno un livello di
similarità tra loro del 98%. Dall’analisi le accessioni 5 e 7 (rispettivamente biotipo Cilento e
Vulture) mostrano un livello di similarità inferiore alle altre. Inoltre ciascun individuo è distinto dal
punto di vista molecolare a differenza di quanto visto con l’analisi ampelografica.
DISCUSSIONI
La storia millenaria della vite e la sua propagazione per via vegetativa hanno prodotto nel
corso dei secoli una proliferazione di popolazioni clonali nelle quali si sono accumulate mutazioni
somatiche di vario genere. Inoltre, la presenza di un così elevato numero di varietà ha provocato una
non corretta identificazione delle stesse, che spesso sono erroneamente definite in base a
denominazioni locali, creando casi di sinonimie e omonimie. Questo quadro così confuso risulta
ulteriormente complicato dai metodi di caratterizzazione varietale, per molto tempo basati
esclusivamente sulla compilazione di schede ampelografiche. Un esempio di quanto appena
affermato, con molta probabilità, riguarda la denominazione e registrazione della cultivar Aglianico
con due differenti appellativi, Aglianico n. e Aglianico del Vulture n., che in realtà identificano la
medesima varietà stando alle metodiche molecolari ad oggi utilizzate per la discriminazione
varietale (SSR). Le differenze rilevate a livello morfologico riguardano foglie adulte e grappoli, e
con buona probabilità sono determinate da fattori ambientali, tuttavia non è da escludere che
essendo questa una varietà di antica origine vi sia anche una variabilità genetica non evidenziabile
con le metodiche molecolari utilizzate in questa ricerca. Rimane quindi da stabilire se i cloni
6appartenenti alla medesima varietà con diversa manifestazione fenotipica ma stesso pattern
molecolare (AFLP e SSR) possano essere considerati appartenenti alla medesima varietà o possano
costituire una nuova varietà. Pelsy et al. (2010) nel loro lavoro riportano che tradizionalmente i
cloni sono attribuiti ad una varietà sulla base di alcuni caratteri comuni ampelografici, tuttavia
piccole differenze a livello fenotipico tra cloni o accessioni della stessa varietà sono tollerate e
accettate. Appare evidente però che i dati morfologici o ampelografici, seppur meno soggettivi
rispetto al passato (UPOV, Bioversity, OIV, 2002), possono avere un ruolo importante se di
supporto ad analisi molecolari, in quanto vi è sempre una certa soggettività nell’analisi. Lavori
analoghi, condotti su accessioni di “Sangiovese” (Scalabrelli et al., 2001), “Parmak” e “Moscato”
(Ergul et al., 2006), hanno riscontrato anch’essi ampia variabilità morfologica tra cloni ascrivibili a
queste varietà e poca variabilità a livello molecolare. Risultati simili sono stati ottenuti anche nello
studio di cloni e mutanti spontanei della cv. “Italia” nel lavoro di Fanizza et al., 2003.
Le analisi molecolari possono essere uno strumento valido per risolvere ogni dubbio di
interpretazione, tuttavia bisognerebbe determinare metodiche capaci di individuare in singole
analisi un numero elevato di polimorfismi. Gli SSR sono sicuramente uno strumento con alte
capacità discriminanti, tuttavia il numero di tali marcatori proposti dal Genres CT96 N° 81potrebbe
non essere sufficiente ad evidenziare differenze cloni. Chiaro è il caso del Pinot che attraverso i
marcatori SSR mostra un identico profilo molecolare per le varietà a bacca bianca, grigia e nera.
Quando non si rilevano differenze tra i campioni esaminati con i marcatori SSR, è
necessario esaminare un elevato numero di loci che coprano in maniera più esaustiva differenti
punti del genoma. In questo caso i marcatori AFLP risultano utili quando si analizza una variabilità
esistente tra presunti cloni della stessa cultivar o tra “cosiddette popolazioni clonali”.
Analogamente a quanto praticato da altri autori (Imazio et al. 2002; Vignani et al., 2002;
Stenkamp et al. 2009) su importanti vitigni quali Pinot, Sangiovese e Traminer, l’analisi con i
marcatori SSR, ampiamente validata nella caratterizzazione varietale (Morgante e Olivieri 1993;
Silvestroni et al. 1997; Sefc et al. 1998 e 2000;) è stata affiancata dall’analisi AFLP. Questi
marcatori all’interno della cultivar Aglianico hanno evidenziato una variabilità molecolare, seppur
minima (in media del 3%), rispetto agli SSR, tuttavia non tale da giustificare l’esistenza di due
diverse cultivar afferenti all’Aglianico. A tale proposito Cervera et al. (1998) avevano proposto che
genotipi di vite con una similarità genetica superiore al 90% potessero essere attribuiti alla
medesima varietà, a condizione che vi sia un sufficiente numero di marcatori molecolari. A tal
proposito risulta importante il lavoro di Fanizza et al., 2003 nel quale viene preso in considerazione
il coefficiente di variazione e viene dimostrato come questo cambi in funzione del numero di bande
analizzate. Un numero di circa 400 bande totali riduce il margine di errore, all’incirca del 5%,
consentendo un più accurato studio di similarità.
Non è importante solo il numero di bande ma anche il numero di caratteri morfologici esaminati in
quanto la similarità genetica misurata dall’analisi con i marcatori AFLP tende a combaciare con i
dati morfologici solo se il numero di caratteri considerati è alto, altrimenti la corrispondenza tende
ad affievolirsi.
Rimane, quindi, la necessità di stabilire e standardizzare nei diversi laboratori di ricerca una
metodica AFLP che, attraverso l’utilizzo: degli stessi enzimi di restrizione, stesse combinazioni di
primer, adeguato numero di bande totali visualizzate e utilizzo di software con stessi settaggi di
lettura; permetta di ottenere dei profili molecolari comparabili, utilizzabili per la definizione di
range di similarità per la discriminazione di cloni e varietà. Inoltre la possibilità di utilizzare, come
avviene per gli SSR, un protocollo di riferimento o comunque delle linee guida, consentirebbe di
7creare dei profili molecolari comparabili da poter utilizzare per creare una banca dati AFLP di
riferimento.
Per dimostrare che la metodica AFLP discussa può essere applicata per la discriminazione di
qualsiasi clone all’interno di una varietà o tra le varietà, sarebbe importante caratterizzare come già
visto con le accessioni di Aglianico, cloni o accessioni appartenenti ad una varietà di recente
costituzione in cui il livello di variabilità molecolare è minore.
8Bibliografia
Barth, S.; Forneck, A.; Verzeletti, F.; Blaich, R; Schumann, F.; 2009: Genotypes and phenotypes of
an ex situ Vitis vinifera ssp sylvestris (Gmel) Beger germplasm collection from the Upper Rhine
Valley. Genet. Resour. Crop Evol. 56, 1171–1181.
Bertsch, C.; Kieffer, F.; Triouleyre, C.; Butterlin, G.; Merdinoglu, D.; Walter, B.; 2003: Molecular
profiling of Vitis vinifera Chardonnays obtained by somatic embryogenesis. J. Int. Sci. Vigne Vin.
37, 223-227.
Blaich, R.; Konradi, J.; Rühl E.; Forneck A.; 2007: Assessing genetic variation among Pinot noir
(Vitis vinifera L) clones with AFLP markers. Am. J. Enol. Vitic. 58 (4), 526–529.
Bonin, A.; Bellemain, E.; Bronken Eidesen, P.; Pompanon, F.; Brochmann, C.; Taberlet, P.; 2004:
How to track and assess genotyping errors in population genetics studies. Mol. Ecol. 13 (11), 3261-
3273.
Bowers, J.E.; Dangl, G.S.; Meredith, C.P.; 1999: Development and characterization of additional
microsatellite DNA markers for grape. Am. J. Enol. Vitic. 50, 243-246.
Catalano, V.; Cersosimo, A.; Stramaglia L.; 1989: Aglianico del Vulture. In: Principali vitigni da
vino coltivati in Italia. Ministero dell’Agricoltura e delle Foreste. ROMA. Vol. I, 145-159.
Crespan, M.; 2003: Evidence on the evolution of polymorphism of microsatellite markers in
varieties of Vitis vinifera L.. Theor. Appl. Genet. 108, 231-237.
Crespan, M.; Milani, N.; 2001: The Muscats: a molecular analysis of synonims, homonyms and
genetic relationships within a large family of grapevine cultivars. Vitis 40, 23-30.
Gonzàles-Techera, A.; Jubany, S.; Ponce de Lèon, I.; Boido, E.; Dellacassa, E.; Carrau, F.M.;
Hinrichsen, P.; Gaggero, C.; 2004: Molecular diversity within clones of cv Tannat (Vitis vinifera).
Vitis 43, 179-185.
Hansen, M.; Kraft, T.; Christiansson, M.; 1999: Evaluation of AFLP in Beta. Theor. Appl. Genet.
98, 845–852.
Hocquigny, S.; Pelsy, F.; Dumas, V.; Kindt, S.; Heloir, M.C.; Merdinoglu, D.; 2004: Diversification
within grapevine cultivars goes through chimeric states. Genome 47, 579-589.
Ibànez, J.; De Andrès, M.T.; Molino, A.; Borrego, J.; 2003: Genetic study of key Spanish grapevine
varieties using microsatellite analysis. Am. J. Enol. Vitic. 54, 22-30.
Imazio, S.; Labra, M.; Grassi, F.; Winfield, M.; Bardini, M.; Scienza, A.; 2002: Molecular tools for
clone identification: the case of the grapevine cultivar “Traminer”. Plant Breeding 121, 531–535.
9Innan, H.; Terauchi, R.; Kahl, G.; Tajima, F.; 1999: A method for estimating nucleotide diversity
from AFLP data. Genetics 151, 1157-1164.
Jahnke, G.; Màjer, J.; Lakatos, A.; Györffyné Molnàr, J.; Deàk, E.; Stefanovits-Bànyai, E.; Varga,
P.; 2009: Isoenzyme and microsatellite analysis of Vitis vinifera L. varieties from the Hungarian
grape germplasm. Sci. Hort. 120, 213–221.
Kimura, M.; Crow, J.F.; 1964: The number of alleles that can be maintained in a finite population.
Genetics 49, 725-738.
Labra, M.; Imazio, S.; Grassi, F.; Rossoni, M.; Sala, F.; 2004: Vine-1 retrotransposon-based
sequence-specific amplified polymorphism for Vitis vinifera L genotyping. Plant Breeding 123,
180-185.
Moio, L.; 2004: Colori, odori ed enologia dell’Aglianico. Regione Campania, Assessorato
Agricoltura.
Nei, M.; 1973: Analysis of gene diversity in subdivided populations. Proc. Natl. Acad. Sci. USA 70,
3321–3323.
Nei, M.; Li, W.H.; 1979: Mathematical model for studying genetic variation in terms of restriction
endonucleases. Proc. Natl. Acad. Sci. USA 76, 5273-5296.
Pelsy, F.; Hocquigny, S.; Moncada, X.; Barbeau, G.; Forget, D.; Hinrichsen, P.; Merdinoglu, D.;
2010: An extensive study of the genetic diversity within seven French wine grape variety
collections. Theor. Appl. Genet. 120, 1219-1231.
Regner, F.; Stadlbauer, A.; Eisenheld, C; Kaserer, H.; 2000: Genetic relationships among Pinots
and related cultivars. Am. J. Enol. Vitic. 51, 7-14.
Riaz, S.; Garrison, K.E.; Dangl, G.S.; Boursiquot, J.M; Meredith, C.P.; 2002. Genetic divergence
and chimerism within ancient asexually propagated winegrape cultivars. J. Am. Soc. Hortic. Sci.
127, 508-514.
Rotondi, A.; Magli, M.; Riccioloni, C.; Baldoni, L.; 2003: Morphological and molecular analyses
for the characterization of a group of Italian olive cultivars. Euphytica 132, 129–137.
Stenkamp, S.H.G.; Becker, M.S.; Hill, B.H.E.; Blaich, R.; Forneck, A.; 2009: Clonal variation and
stability assay of chimeric Pinot Meunier (Vitis vinifera L) and descending sports. Euphytica 165,
197–209.
This, P.; Jung, A.; Boccacci, P.; Borrego, J.; Botta, R.; Costantini, L.; Crespan, M.; Dangl, G.S.;
Eisenheld, C.; Ferreira-Monteiro, F.; Grando, M.S.; Ibanez, J.; Lacombe, T.; Laucou, V.;
Magalhaes, R.; Meredith, C.P.; Milani, N.; Peterlunger, E.; Regner, F.; Zurlini, L.; Maul, E.; 2004:
10Development of a standard set of microsatellite references alleles for identification of grape
cultivars. Theor. Appl. Genet. 109, 1448-1458.
Upov; Bioversity; Oiv; 2002: OIV descriptor list for grape varieties and Vitis species. 2nd edition
Organisation Internationale de la Vigne et du Vin, Paris, France/International Plant Genetic
Resources Institute, Rome.
Vignani, R.; Scali, M.; Masi, E.; 2002: Genomic variability in Vitis vinifera L “Sangiovese”
assessed by microsatellite and non-radioactive AFLP test. Electronic Journal of Biotechnology 5 (1),
1-11. Available on line at http://www.ejborg/content/vol5/issue1/full/2.
Violante, C.; Bordignon, S.; 1962: Principali vitigni da vino coltivati in Italia. Ministero delle
Politiche Agricole e Forestali. ROMA. Vol. III, 1-10.
Vos, P.; Hogers, R.; Bleeker, M.; Reijans, M.; van de Lee, T.; Hornes, M.; Frijters, A.; Pot, J.;
Peleman, J.; Kuiper, M.; Zabeau, M.; 1995: AFLP: a new technique for DNA fingerprinting. Nucl.
Acids Res. 23, 4407-4414.
Zoghlami, N.; Riahi, L.; Laucou, V.; Lacombe, T.; Mliki, A.; Ghorbel, A.; This, P.; 2009: Origin
and genetic diversity of Tunisian grapes as revealed by microsatellite markers. Sci. Hort. 120, 479–
486.
11Tab. 1: Caratterizzazione ampelografica, mediante l’utilizzo di 14 descrittori primari OIV (2ND Edition 2007), di 2 cloni certificati
di Aglianico n. e Aglianico del Vulture e di 31 accessioni ad essi ascrivibili. I biotipi Sannio, Taurasi e Cilento appartengono
alla varietà Aglianico n.. I dati riportati riguardano gli anni 2007, 2008 e 2009. In rosso sono riportati i valori delle accessioni
che differiscono rispetto ai due cloni certificati. In grassetto sono evidenziate le differenze tra i due cloni utilizzati come
riferimento.
germogli acini
foglie descrittori
descrittori descrittori
n°
Biotipo 001 004 016 051 067 068 070 076 079 081-2 084 087 223 225
acces.
1 Sannio 5 3 1 1 4 2-3 1 3 5 1 5 3 2 6
2 Vulture 5 3 1 1 4 2-3 1 3 5 1 5 3 2 6
3 Vulture 5 3 1 1 4 2-3 1 3 5 1 5 3 2 6
4 Taurasi 5 5 1 1 2 2 2 5 3 1 5 3 2 6
5 Cilento 5 5 1 1 4 2 1 3 7 1 7 5 2 6
6 Cilento 5 5 1 1 4 2 1 3 7 1 7 5 2 6
7 Vulture 5 3 1 1 2 3 1 5 7 1 7 7 2 6
8 Vulture 5 5 1 1 2 2 2 5 3 1 5 3 2 6
9 Taurasi 5 5 1 1 3 3 1 2 5 1 5 3 2 6
10 Vulture 5 5 1 1 3 3 1 2 5-7 1 5 3 2 6
11 Vulture 5 5 1 1 3 3 1 2 5 1 5 5 2 6
12 Vulture 5 5 1 1 3-4 3 1 2 5 1 5 5 2 6
13 Taurasi 5 5 1 1 3-4 3 1 2 5 1 5 5 2 6
14 Vulture 5 5 1 1 3 2 1 2 5 1 5 5 2 6
15 Taurasi 5 5 1 1 3 3 1 2 3 1 5-7 5 2 6
16 Taurasi 5 5 1 1 3 3 1 2 7 1 3 3 2 6
17 Taurasi 5 5 1 1 3 2 1 2 5 1 3 5 2 6
18 Taurasi 5 5 1 1 3 2 1 2 3 1 3 3 2 6
19 Taurasi 5 5 1 1 3 2 1 2 3 1 5 5 2 6
20 Vulture 5 5 1 1 3 2 1 2 5 1 3 3 2 6
21 Taurasi 5 5 1 1 3 3 1 2 5 1 3 3 2 6
22 Taurasi 5 5 1 1 3 2 1 2 3 1 3 3 2 6
23 Taurasi 5 5 1 1 3 3 1 2 3-5 1 3-5 3 2 6
24 Taurasi 5 5 1 1 2 2 1 2 5 1 3 3 2 6
25 Taurasi 5 5 1 1 4 3 1 2 3-5 1 3 3 2 6
26 Taurasi 5 5 1 1 3 3 1 2 5 1 5 3 2 6
27 Taurasi 5 5 1 1 3 2 1 2 5 1 5 3 2 6
28 Taurasi 5 5 1 1 4 2 1 3 7 1 7 5 2 6
29 Taurasi 5 5 1 1 2 2 1 2 5 1 5 3 2 6
30 Taurasi 5 5 1 1 3 2 1 2 3 1 5 3 2 6
31 Cilento 5 5 1 1 3 3 1 2 5 1 3 3 2 6
Aglianico
5 5 1 1 3 3 1 2 5-7 1 3 3 2 6
n.
Aglianico
5 5 1 1 4 2 1 3 7 1 7 5 2 6
Vult.
12Tab. 2: Caratterizzazione ampelografica, mediante l’utilizzo di 16 descrittori secondari OIV (2ND Edition 2007), di 2 cloni
certificati di Aglianico n. e Aglianico del Vulture e di 31 accessioni ad essi ascrivibili. I biotipi Sannio, Taurasi e Cilento
appartengono alla varietà Aglianico n.. I dati riportati riguardano gli anni 2007, 2008 e 2009.
germogli foglie
grappoli descrittori acini descrittori
descrittore descrittori
n° 083-
Biotipo 003 065 074 078 080 081-1 202 203 204 206 208 209 231 235 241
acces. 2
1 Sannio 1 5 3 9 9 7 7 5-7 3 2 2 1 1 3
3 5
2 Vulture 1 5 3 9 9 5-7 3-5 5 3-5 2 1-2 1 1 3
1 5
3 Vulture 1 5 3 9 9 3-5 3-5 5 3-5 2 1-2 1 1 3
1 5
4 Taurasi 2 3 1 1 1 5-7 5-7 7 3 2 2 1 1 3
1 5
5 Cilento 1 3 3 1 1 7 5-7 5 3-5 2 2 1 1 3
1 7
6 Cilento 1 3 3 1 1 5-7 3-5 7 3-5 2 2 1 1 3
1 7
7 Vulture 1 5 3 1 1 1-3 3 3 5-7 1 2 1 1 3
1 5
8 Vulture 2 3 1 1 1 5 5 3-5 3 2 2 1 1 3
1 5
9 Taurasi 1 5 1 1 9 5 3 5 3 2 2 1 1 3
1 5
10 Vulture 2 3 1 1 1 1 1-3 5 3 1-2 1 1 1 3
1 5
11 Vulture 1 5 1 1 9 7 5 5 3 2 2 1 1 3
1 5
12 Vulture
1 5 1-2 5 2 1 9 7 7 5 3 2 2 1 1 3
13 Taurasi 1 5 1 1 9 5 3 5 3 2 1 1 1 3
1 5
14 Vulture 1 5 1-2 1 9 7 3-5 5-7 1-3 1 1 1 1 3
1 5
15 Taurasi 1 3 3 1 1 5 1-3 5 3 2 1 1 1 3
1 5
16 Taurasi
1 5-7 1 5 1 9 9 5 3 7 3 2 1 1 1 3
17 Taurasi 1 5 1 1 1 5-7 3-5 3 5 2 2 1 1 3
1 5
18 Taurasi 2 5 2 1 9 7 5 5 7 2 2 1 1 3
1 5
19 Taurasi 1 5 2 1 9 5 3 5 3 2 1 1 1 3
1 5
20 Vulture 1 5 1 1 1 5 3-5 5-7 5 2 2 1 1 3
1 5
21 Taurasi 1 5-7 1 1 1 7 5-7 3 5 2 2 1 1 3
1 5
22 Taurasi 1 5 1 1 1 3-5 3 3-5 5 1 2 1 1 3
1 5
23 Taurasi 1 5 1 1 9 3-5 3-5 5 3 2 2 1 1 3
1 5
24 Taurasi
1 5 1-2 5 1 1 1 7 5 5 5 2 2 1 1 3
25 Taurasi 1 3 1 1 1 5-7 5-7 5-7 5 2 2 1 1 3
1 5
26 Taurasi 1 5 1 1 9 5 3 5 3 2 2 1 1 3
1 5
27 Taurasi 1 5 1 1 1 5-7 5 5 3 2 2 1 1 3
1 5
28 Taurasi 1 3 3 1 1 5-7 3-5 7 3-5 2 2 1 1 3
1 7
29 Taurasi 1 3-5 1 1 9 7 5 3 7 2 2 1 1 3
1 5
30 Taurasi 1 5 1 9 9 5 3 3 5 1 2 1 1 3
1 5
31 Cilento 1 5 1 1 1 7 5-7 3-5 5 2 2 1 1 3
1 5
Aglianico
1 5 1 5 1 1 1 3 3 3 3 1 1 1 1 3
n.
Aglianico
1 7 1 3 3 1 1 5 5 5-7 5 2 2 1 1 3
Vult.
.
13Fig. 1: Dendrogramma di similarità costruito attraverso il software NTSYS, utilizzando il coefficiente di distribuzione DIST e
l’algoritmo UPGMA. Si formano complessivamente 2 cluster principali e 4 sotto-cluster. Nel terzo sottocluster si raggruppano le due
cultivar certificate mentre il quarto sotto-cluster è costituito dalla sola accessione numero 7 biotipo Vulture. Non è possibile da questa
analisi distinguere tutte le accessioni dal punto di vista morfologico. In rosso sono riportate le accessioni ascrivibili all’Aglianico del
Vulture, in nero le accessioni ascrivibili all’Aglianico n.; mentre in grassetto e sottolineate sono evidenziati i cloni di riferimento
14Tab. 3:Lettura dei profili molecolari ottenuti con 13 combinazioni di
marcatori SSR. Con l’asterisco sono indicati i marcatori proposti dal EU
project Genres CT96 N° 81
SSR marker Allele size (bp)
ISV3 134 146
ISV4 171 179
VVMD28 229 259
VMCNG4b9 164 168
VrZAG79* 244 246
ISV2 144 166
VrZAG62* 187 187
VVMD7* 238 238
VVS2* 151 155
VVMD27* 182 188
VVMD5* 231 245
VVMD25 249 263
VVMD32 250 256
15Fig. 2: Profilo AFLP letto mediante utilizzo di un free Software: Genographer. Il software
permette di trasformare i picchi degli elettroferogrammi in un’immagine virtuale di un gel. Le
frecce in nero sulla sinistra permettono di evidenziare alcuni polimorfismi presenti nella
combinazione E44-M59. Solo le bande al di sopra delle 100bp sono state considerate nel
lavoro.
16Tab. 4:Lettura dei profili molecolari ottenuti con 10 combinazioni di marcatori AFLP. La combinazione E43-M48 ha mostrato il maggior numero di bande totali e di bande
polimorfiche. La combinazione E44-M61 ha invece mostrato il più alto rapporto, espresso in percentuale, tra bande amplificate e bande polimorfiche.
Combinazioni Bande totali Bande polimorfiche Alleli effettivi Gene diversity Polymorphic Average proportion Nucleotide
di primer amplificate information content of shared bands diversity
n. n. % Ne GD PIC % π (x 1000)
E43-M48 61 19 31.1 1.167 0.105 0.043 98.1 0.959
E43-M50 27 4 14.8 1.087 0.053 0.022 99.0 0.604
E43-M51 45 16 35.6 1.189 0.120 0.150 98.0 0.533
E44-M48 34 11 32.4 1.185 0.118 0.079 97.4 0.492
E44-M59 36 12 33.3 1.242 0.142 0.074 96.7 0.995
E44-M61 31 12 38.7 1.219 0.136 0.050 97.7 1.107
E45-M61 30 11 36.7 1.198 0.126 0.105 96.7 1.281
E46-M49 25 7 28.0 1.185 0.110 0.047 97.8 1.673
E46-M59 53 8 15.1 1.087 0.054 0.034 98.8 1.179
E46-M61 35 8 22.9 1.118 0.077 0.022 98.9 1.703
Total 377 108 1.005
Mean 37.7 10.8 28.9 1.168 0.110 0.063 97.9 1.053
SD 11.71 4.39 0.09 0.05 0.04 0.04 0.01 0.43
17Fig. 3: Dendrogramma di similarità costruito sulla base dei dati molecolari AFLP, utilizzando il Software NTSYS con coefficiente di
similarità DICE e algoritmo UPGMA. Le accessioni mostrano complessivamente una similarità maggiore del 94%. L’Aglianico n. e
L’Aglianico del Vulture condividono una similarità genetica del 98%. Inoltre tutti gli individui sono distinguibili, mostrando una
diversità, seppur minima, a livello molecolare.
18Puoi anche leggere

















































