Il Gallo forcello in AMBIENTE ALPINO - UNCZA
←
→
Trascrizione del contenuto della pagina
Se il tuo browser non visualizza correttamente la pagina, ti preghiamo di leggere il contenuto della pagina quaggiù
[ricerca scientifica]
Il Gallo forcello
in AMBIENTE ALPINO
Messa a punto di un importanza l’attivazione di un accordo di colla- BARBARA
CRESTANELLO
borazione per la raccolta di campioni di fauna
protocollo genomico Department of
tipica alpina, idonei all’analisi genetica, secondo Biodiversity and
per l’acquisizione un protocollo comune condiviso a livello delle Molecular Ecology
Research and
di dati finalizzati Alpi italiane. Lo scopo è la creazione di una ban- Innovation Centre -
Fondazione Edmund
alla gestione, in una ca dati biologica che sarà base fondamentale per
Mach
la realizzazione di attività di studio e ricerca in
ricerca UNCZA
ambito genetico molecolare. In collaborazione con
ALICE FRASER
Ed è proprio su parte di questi campioni dottoranda FEM-UNIFE
che si basa la recente collaborazione tra UNCZA
e FEM che aveva come obiettivo la messa a punto
di un protocollo innovativo, idoneo all’acquisizione
INTRODUZIONE di nuovi dati applicabili alla gestione e conserva-
Il gruppo di Genetica della Conservazione della zione del Gallo forcello in territorio alpino italia-
Fondazione Edmund Mach (FEM) è da più anni no. Lo studio della variabilità genetica permette di
impegnato nella caratterizzazione genetica di indagare gli effetti degli eventi passati, valutare le
diverse specie di fauna alpina con scopi di con- dinamiche future e prevedere gli esiti delle azioni
servazione, gestione e/o monitoraggio. Aspetto dell’uomo e dei cambiamenti ambientali nelle po-
fondamentale per la buona riuscita di questi polazioni naturali.
progetti è la realizzazione di un campionamento La variabilità genetica costituisce il livel-
ampio ed esaustivo, che permetta di raccogliere lo strutturale più semplice della biodiversità
una serie di dati in grado di fornire un quadro il complessiva di un ecosistema ed è lo strumento
più possibile completo della situazione attuale. necessario alle popolazioni per evolvere ed af-
Per questo UNCZA, insieme a FEM e ISPRA frontare i cambiamenti ambientali che negli ul-
(Istituto Superiore per la Protezione e la Ricer- timi decenni avvengono in maniera sempre più
ca Ambientale), hanno ritenuto di fondamentale rapida.
4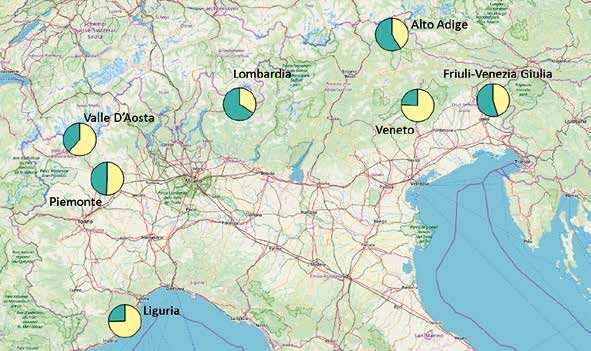
Gli effetti negativi dell’impoverimento gene- tetraonidi del Paleartico con una estensione lon-
tico sulla fitness e sulle capacità di adattamento e gitudinale totale che supera i 6000 km. La distri-
sopravvivenza delle popolazioni naturali sono sta- buzione di questa specie è abbastanza continua
ti ormai ampiamente dimostrati: di conseguenza, nella parte nord-orientale dell’areale, grazie alla
le politiche di conservazione, gestione o uso so- disponibilità di un habitat pressoché ininterrot-
stenibile delle risorse faunistiche di un territorio, to costituito dalle foreste boreali, mentre risulta
soprattutto quelle soggette a forte sfruttamento piuttosto frammentata nella porzione sud-occi-
(caccia, pesca, traslocazioni, ecc.), non possono dentale, in particolar modo in Europa centrale,
più trascurare la salvaguardia o il ripristino della dove è presente in piccoli nuclei isolati, relitti dei
biodiversità a questo livello. fenomeni glaciali.
Nel presente lavoro sono stati applicati i In Italia il Fagiano di monte è presente sui
metodi di una nuova disciplina, conosciuta con il rilievi alpini e prealpini, a quote comprese tra i
nome di “genomica di conservazione”, che utilizza 900 ed i 2000 m s.l.m.
le informazioni ottenute dallo studio della variabi- Le popolazioni di Gallo forcello sono con-
lità presente nell’intero genoma al fine di preserva- siderate pressoché stabili in gran parte del loro
re la vitalità e la biodiversità di una specie. Questo areale salvo che in Europa centrale ed occiden-
nuovo approccio permetterà di studiare, con un tale, dove i contingenti della specie hanno subi-
livello di dettaglio mai raggiunto prima, aspetti im- to a partire dagli anni ’70 una rapida contrazio-
portanti legati alla storia demografica, all’ecologia ne, che ha portato fino ad eventi di estinzione
e alla biologia di questa specie. locale. Nell’Europa centrale, la popolazione più
Il Gallo forcello è una specie a corologia eu- ampia e relativamente stabile è distribuita lungo
rosibirica boreoalpina. Il suo areale distributivo, la catena alpina.
compreso fra le medie e le alte latitudini del con- Il pericolo maggiore che la conservazione
Tabella 1: Numero di campioni di fauna tipica alpina raccolti, nelle Alpi Italiane e Svizzere,
nell’ambito dell’accordo di collaborazione tra UNCZA, FEM forcello
tinente euroasiatico, è il più vasto fra quelli dei del Gallo devea fronteggiare
ed ISPRA a livello
partire dalla glo-
stagione
venatoria 2010/2011. I campioni sono stati suddivisi per Regione e provincia di campionamento.
Tabella 1. K6"+$"
Numero di campioni di !"#$%&"'()%*$&+$, -,#$,&%./$.0%&1"
Gallo forcello 2%13)&$+" 2"/)%&" (")&$+" -),&+%4$&% 5"6)".7$,&+,
fauna tipica alpina raccolti,
-)$34$89"&":$,.;$34$, < 8 8 8 8 8
nelle Alpi Italiane e Svizzere,
!"#$%&"&% ' ( ( ( ( (
nell’ambito dell’accordo di
)$*&% + ( ( ( ( (
collaborazione tra UNCZA,
5$#3)$, = 8 8 8 8 8
FEM ed ISPRA a partire
,-.%#*/ + ( ( ( ( (
dalla stagione venatoria
5%07,)/$, >>= ?@A 8 BC 8 =
2010/2011. I campioni sono
0%#1/-" 23 4 ( ( ( (
stati suddivisi per regione e
0#%56*/ '+ + ( 2 ( (
provincia di campionamento.
7"-" '89 29: ( ; ( (
?E 8 CD 8 C
0*%>>/ + ( ( ( ( (
!
7A&%" '8 2 ( 3 ( (
B"#*&" ' C ( : ( (
@%#D/&"(7A5*"(E55">/ 8+ '; ( ; ( ;
@%#6%>>* C8 ;+ ( 2 ( (
F)"&1$&%.,41%.,/$#" DC> D= E@ >E > DD
0">F/&" C4 ' 3' 23 ' 3
B#%&G" 2: '8 ': ; ; 4
9,44".GHI%J1, AE CA 8 DA 8 >?
H"5G/ C: ;C ( 'C ( 24
9"&"1% CD C < 8 8 8
0%>>A&" 2 ' ( ( ( (
@*6%&F/ 'C ' 3 ( ( (
K*$::"), D= 8 8 B 8 8
7/&G"&I@/>>%5% '+ ( ( 8 ( (
F%1,4".+%064"JJ$*% E< D>B > B>
Tabella 2: Numero di campioni di fagiano di monte analizzati nel presente progetto, suddivisi 5
per Regione e Provincia. Per ciascuna Regione è riportata l’eterozigosità media attesa (HE),
calcolata usando il pacchetto 4P (Benazzo et al., 2015). 
Tabella 1: Numero di campioni di fauna tipica alpina raccolti, nelle Alpi Italiane e Svizzere,
nell’ambito
bale è dell’accordo di collaborazione
l’alterazione dell’habitat. Il degrado tra
e laUNCZA, FEM ed
RISULTATI ISPRA a partire dalla stagione
E CONCLUSIONI
venatoria 2010/2011. I campioni sono stati suddivisi per Regione e provincia di campionamento.
frammentazione degli habitat hanno isolato certe Dall’analisi dei 173 campioni di Fagiano di monte
popolazioni, rendendole particolarmente esposte con ilK6"+$"
metodo GBS sono stati ottenuti un totale di
al rischio di estinzione.
!"#$%&"'()%*$&+$, -,#$,&%./$.0%&1" 2%13)&$+" 2.442 Polimorfismi
2"/)%&" (")&$+" di Singolo Nucleotide5"6)".7$,&+,
-),&+%4$&% (SNP).
-)$34$89"&":$,.;$34$, < 8 8 numero di marcatori
Il 8 ottenuto 8 è buono consi- 8
!"#$%&"&% ' (
METODICA derando la qualità dei campioni originali, ma (in-
( ( (
)$*&% + ( ( ( ( (
Grazie alla concreta collaborazione
5$#3)$, = dell’UNCZA
8 e feriore
8 a quello 8atteso (normalmente8 tra i 10.000
8
dei suoi soci, nell’ambito+ dell’accordo di( collabo-
,-.%#*/ e( i 30.000 SNPs). ( ( (
5%07,)/$,
razione UNCZA-FEM-ISPRA, >>= ?@A raccolti
sono stati 8 Tuttavia,BCun sequenziamento 8 aggiuntivo,=at-
0%#1/-" 23 4 ( ( ( (
più di 1500 campioni (piume,
0#%56*/ '+
pelo, tessuto)
+
appar- tualmente
(
in corso,
2
permetterà( di raggiungere (
i
7"-"tenenti alle 6 specie di'89 fauna tipica alpina
29: presen- risultati
( attesi e;di approfondire( le analisi. (
ti in territorio italiano (Tabella
?E l’analisi
8 della CD
diversità genetica 8 presente traC gli
0*%>>/ I campioni di tessuto + e piume sono( stati (
individui possa (essere fonte di( informazione per (
!estratti tramite il sistema
7A&%" '8
automatico di
2
separa-
(
la
3
gestione di questa specie. (
( (
B"#*&" ' C ( : (
zione magnetica Thermo
@%#D/&"(7A5*"(E55">/ 8+ Scientific '; KingFi-
TM
( I valori di; eterozigosità (attesi e osservati,;
sherTM Flex (Thermo C8
@%#6%>>* Fisher Scientific,
;+ Bonn, calcolati
( per ciascuna
2 regione, ( sono compresi (
F)"&1$&%.,41%.,/$#" DC> D= E@
Germany). nell’intervallo>E >
0,2-0,3. L’eterozigosità DD
è un indi-
0">F/&" C4 ' 3' 23 ' 3
B#%&G" La tipizzazione dei 2: campioni è stata
'8 effet- ':ce che varia tra; 0 e 1 ed indica ; la quantità di 4va-
tuata tramite la metodica
9,44".GHI%J1, AE “GenotypingCA by se- riabilità
8 genetica
DA presente nei8 dati; più il valore >?
H"5G/
quencing” (GBS – Elshire C: ;C che si è
et al. 2011), è( vicino ad 1 'C (
maggiore è la variabilità 24
genetica
9"&"1% CD C < 8 8 8
dimostrata efficace per2 l’identificazione
0%>>A&" '
di un presente.
(
I livelli
(
di eterozigosità(
riscontrati( in
elevato numero di polimorfismi
@*6%&F/ 'C di singolo
' nu- 3questa analisi evidenziano
( livelli
( medio-bassi( di
cleotide (SNPs) in specie
K*$::"), D= non modello. 8 Questa variabilità
8 genetica
B che potrebbero8 essere un 8se-
7/&G"&I@/>>%5% '+ ( (gnale di sofferenza
8 (
della specie studiata. Le rela- (
tecnologia si basa sul sequenziamento NGS di li-
F%1,4".+%064"JJ$*% E< D>B > B>
brerie di DNA genomico, arricchite in sequenze zioni genetiche esistenti tra i diversi individui e
codificanti mediante l’utilizzo di enzimi di restri- regioni sono rappresentate in modo semplificato
zione, e permette il sequenziamento simultaneo
Tabella 2: Numero di campioni di fagiano di monte nelanalizzati
grafico ottenuto dall’analisi progetto,
nel presente delle componen-
suddivisi
per Regione e Provincia. Per ciascuna Regione è riportata l’eterozigosità media attesa la(H ),
di numerosi campioni (96 o 384). ti principali (PCA; Fig. 2). Il grafico evidenzia
E
calcolata usando il pacchetto 4P (Benazzo et al., 2015).
Tabella 2
!"#$%&"' ()%*$&+$,' -' ./' !"#$%&"' ()%*$&+$,! -! ./' Numero di campioni di
! "#$%&'# ! () ! '
! *#+,& ! -. ! ! fagiano di monte analizzati
012%'03$#"
8%2 ! !
9: ;'
4,11"'560%72,
8%2 ! =? ! ;'
nel presente progetto,
suddivisi per regione e
"/01&2# ! 3 ! ' 45'/# ! -( ! ! provincia. Per ciascuna
"0/+67& ! 8 '! ! 9/0:&#;45+7#;? ;'
D)$E1$F4"&"G$,'
H$E1$, ! DB7'/ ! . ! !
"/$$5'# ! @ ! ' 8%2 ! I ! ;
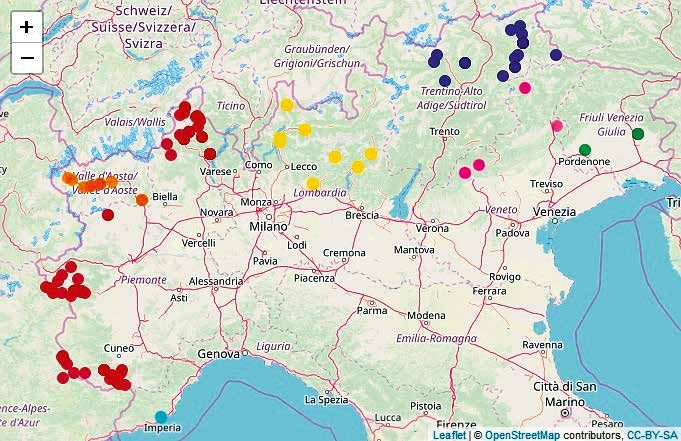
Figura 1.
Siti di campionamento
degli individui di fagiano
di monte utilizzati nel
presente progetto. Ogni
punto rappresenta un
singolo individuo e i colori
rappresentano le Regioni di
campionamento come da
riquadro. Mappa creata in R
(R Core Team, 2018) usando il
pacchetto leaflet (Graul, 2016).
presenza di una moderata struttura geografica. proporzione di appartenenza a ciascuno dei due
Le ellissi, che rappresentano le regioni, sono tra gruppi, sono stati riportati anche sotto forma di
loro in parte sovrapposte e molti individui (rap- grafici a torta sulla mappa del territorio alpino,
presentati dai punti) si raggruppano nella parte considerando la composizione complessiva per
centrale del grafico, indipendentemente dalla ciascuna delle regioni. Analogamente ai risultati
regione di campionamento (rappresentata dai della PCA, non si riscontrano raggruppamenti su
colori). Inoltre, piuttosto che il raggruppamen- base geografica.
to di individui sulla base della provenienza ge- Il differenziamento degli individui in popo-
ografica, alcune regioni (Piemonte, Lombardia lazioni può essere il risultato della distanza geo-
e Alto-Adige), mostrano un’alta differenziazione grafica che li separa ed essere, ad esempio, lega-
al loro interno con individui che si espandono to alle capacità di dispersione di una specie. Per
sui grafici lungo entrambi gli assi. testare se la differenziazione genetica osservata
Per identificare la struttura genetica più nel fagiano di monte sia legata alla distanza geo-
compatibile con i nostri dati è stato applicato un grafica è stato utilizzato il metodo di ‘isolamento
apposito algoritmo che ha dato come risultato che per distanza’ (IBD), che studia come cambiano
gli individui analizzati possono essere ricondotti a le distanze genetiche al variare delle distanze ge-
due gruppi geneticamente distinti. ografiche. Se i dati seguono un modello di isola-
Nel grafico (Figura 3) gli individui rap- mento per distanza, maggiore è la distanza geo-
presentati da una barra composta da due colori grafica che separa due individui maggiore sarà la
sono individui ibridi tra i due gruppi genetici loro distanza genetica.
individuati (POP1 verde, POP2 giallo). La mag- I dati mostrano un isolamento per distanza
gior parte degli individui studiati presenta una significativo (0,33, p=0,0012).
componente genetica mista. Per valutare meglio In generale, i dati genetici attualmente
la distribuzione geografica dei due gruppi gene- disponibili sono solo moderatamente influen-
tici emersi dalle analisi, i risultati, in termini di zati dai pattern geografici. Non è stata, infatti,
7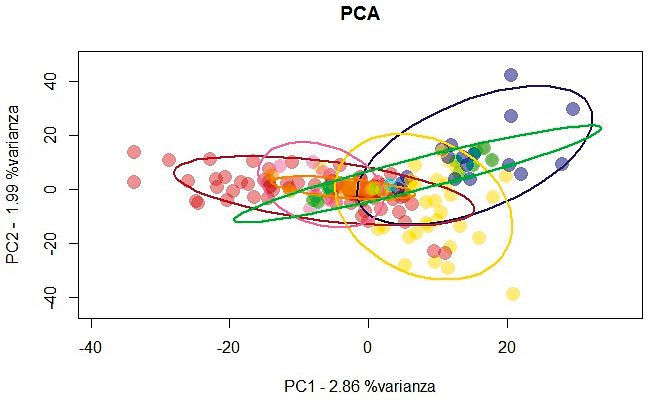
Figura 2.
Analisi delle Component
Principali (PCA). Ogni
punto rappresenta un
individuo e le ellissi
rappresentano le regioni
(90% intervallo di confidenza
dal centroide), i colori
rappresentano le regioni
di campionamento come
da Figura 1. Più vicini sono
due punti sul grafico più
simili geneticamente sono
i corrispondenti individui.
Analisi effettuata in R (R
Core Team, 2018) usando
il pacchetto adegenet
(Jombart et al., 2008).
evidenziata la presenza di popolazioni geneti- una specie adattata all’ambiente di bassa mon-
che geograficamente distinte, ma solo segnali tagna e con distanze di dispersione post-natale
significativi di isolamento per distanza. Questo limitate, le catene montuose o le ampie val-
sta ad indicare un aumento progressivo della li presenti lungo l’Arco Alpino non sembrano
diversità genetica all’aumentare della distanza essere delle reali barriere alla dispersione per
geografica. Diversamente da quanto atteso, per questa specie.
8Figura 3.
Contributo genetico delle 2
popolazioni inferite (POP1
verde, POP2 giallo) nelle
Regioni e nei genotipi
individuali. Nella mappa
ogni cerchio rappresenta una
regione, nel grafico ogni barra
rappresenta un individuo e
gli individui sono raggruppati
per regione di provenienza
(colori come da Figura 1).
Mappa creata con QGIS (QGIS
Development Team, 2019).
Rimane da approfondire la presenza, sull’Ar- principali attraverso le quali hanno luogo la disper-
co Alpino, di due gruppi geneticamente distinti, sione degli individui ed il flusso genico. Infatti, seb-
seppure omogeneamente distribuiti. La distri- bene i dati non abbiano evidenziato fenomeni di
buzione omogenea sul territorio di individui con isolamento genetico, la frammentazione delle aree
componente genetica mista, ma anche di individui idonee per la specie e la suddivisione in un gran
‘puri’ con componente genetica unica (completa- numero di sottopopolazioni, tra loro diversamente
mente gialli o verdi nel grafico), fa pensare a feno- connesse e, soprattutto, con densità demografiche
meni recenti di mescolamento tra gruppi prece- molto variabili, rimangono tra le principali minac-
dentemente isolati, dei quali sarebbe importante ce alla conservazione del Gallo forcello.
identificare la provenienza. Le analisi attualmente in corso, che preve-
In conclusione, una gestione attenta ed effi- dono oltre al sequenziamento aggiuntivo, l’analisi
cace della specie a livello alpino, dovrà tenere pre- di nuove popolazioni distribuite sull’arco alpino
sente il livello medio-basso di variabilità genetica italiano, attraverso una maggiore informazione
riscontrata. Le scelte, in termini di abbondanza e genetica e una più ampia copertura geografica,
distribuzione dei prelievi, dovrebbero essere ridi- consentiranno di valutare più in dettaglio, su sca-
scusse non solo in funzione dei criteri demografi- la più fine, quanto finora riscontrato per fornire
ci ma, anche, identificando e preservando le aree così specifici suggerimenti gestionali.
9Puoi anche leggere

















































