DANNI AL DNA COME CAUSA DELL'INVECCHIAMENTO
←
→
Trascrizione del contenuto della pagina
Se il tuo browser non visualizza correttamente la pagina, ti preghiamo di leggere il contenuto della pagina quaggiù
DANNI AL DNA COME CAUSA
DELL’INVECCHIAMENTO
La Drosophila melanogaster è un organismo modello, ovvero uno degli organismi più
studiati nella ricerca biologica, in particolare nella genetica e nella divulgazione
biologica.
I motivi sono molteplici:
• Si tratta di un insetto piccolo e facile da allevare in laboratorio;
• Ha un breve tempo di generazione (circa 2 settimane) e una elevata
produttività (le femmine possono deporre 400 uova in 10 giorni);
• Le larve mature mostrano cromosomi "giganti" nelle ghiandole salivari;
• Hanno solo 4 paia di cromosomi: 3 autosomi e 1 sessuale;
• I maschi non mostrano ricombinazioni genetiche, facilitando gli studi
genetici;
Da un punto di vista genetico l'uomo e il moscerino della frutta sono abbastanza
simili.
Circa il 60% delle malattie genetiche conosciute si possono verificare nel
patrimonio genetico del moscerino e circa il 50% delle proteine della
Drosophila hanno un analogo nei mammiferi.
La Drosophila viene usata come modello genetico per varie malattie umane,
inclusi i disturbi neurodegenerativi come il morbo di Parkinson e il morbo di
Alzheimer.
Il moscerino viene utilizzato anche per studiare il meccanismo biologico del sistema
immunitario, del diabete, del cancro e persino dell'abuso di sostanze stupefacenti
Michael R. Rose è un professore presso il Dipartimento di Ecologia e Biologia
Evolutiva presso l' Università della California, Irvine . La principale area di lavoro è
stata l' evoluzione dell’invecchiamento, affrontato sia teoricamente che
empiricamente attraverso la tecnica di evoluzione sperimentale .
Nel 1991, ha pubblicato Evolutionary Biology of Aging esplorando una visione del
soggetto basato sulla dipleiotropia antagonista , dove esprime l'ipotesi che
l'invecchiamento è causato da geni che hanno due effetti, uno che agisce presto nella
vita e l'altro molto più tardi.
I geni sono favoriti dalla selezione naturale come risultato dei benefici delle loro
prime fasi di vita, e il prezzo che maturano molto più tardi appaiono come accessori
effetti collaterali che noi identifichiamo come l'invecchiamento. Dr. Rose ha anche
suggerito che l'invecchiamento può fermarsi in una seconda fase della vita.
Il fenomeno è stato descritto da George C. Williams nel 1957, ma fu Rose che ha
coniato la frase " pleiotropia antagonista ". Il laboratorio di Rose ha condotto la più
lunga sperimentazione di selezione artificiale progettato per testare la teoria della
pleiotropia antagonista.
1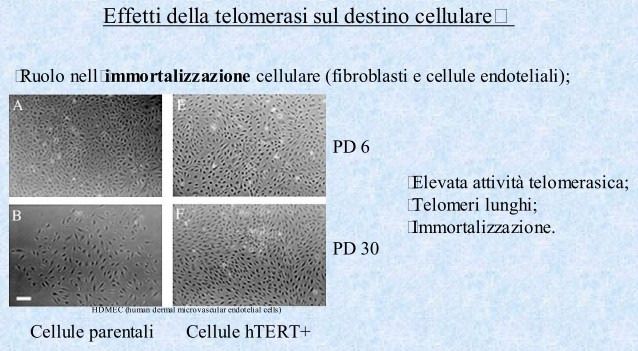
Sperimentazione
Incrociando, quindi, tra di loro maschi e femmine sempre più longevi della
“Drosophila melanogaster”, il moscerino della frutta, Michael R.Rose, è riuscito ad
ottenere una popolazione che è in grado di vivere il doppio delle normali popolazioni
di laboratorio.
La previsione dell'ipotesi pleiotropia antagonista era che queste mosche longevi
avrebbero avuto presto una fertilità molto più basso nella vita. Il risultato è stato il
contrario: le mosche longevi in realtà depongono più uova in ogni fase della vita.
Ad ogni stadio di sviluppo si dimostrano più robusti resistendo meglio allo stress.
Rose spiega questo risultato in termini di una interazione tra genotipo e ambiente. Le
mosche longevi mostrano debolezze che li rendono concorrenti poveri in natura, e,
forse, questi tratti sono i veri settori della pleiotropia antagonista
Il lavoro di Rose sui moscerini della frutta si inserisce in una linea di ricerca più
ampia che si propone di capire come mai gli esseri umani invecchino o come mai il
deperimento e la morte siano per noi processi inevitabili
SCOPO → Che cosa manca ai moscerini normali?
Egli iniziò con il confrontare le proteine sintetizzate dagli insetti dell’esperimento con
quelle prodotte dagli insetti normali. L’enzima superossidodismutasi sembrava essere
più attivo.
Rose è riuscito ad applicare la sua teoria in modo molto convincente, partendo dal
presupposto che l’invecchiamento si attua “a causa di un declino nella forza di
selezione nell’età adulta”.
Selezionando i moscerini longevi è riuscito ad ottenerne alcuni che vivevano 80
giorni contro i normali 25. Tradotti sull’uomo questi numeri significano una
sopravvivenza di almeno 160 anni.
Ma le ricerche di Rose hanno scarsa applicabilità:
a) gli studi devono essere condotti specie per specie. I risultati ottenuti su
una specie non valgono se trasferiti in un’altra;
b) le tecniche di selezione ottengono risultati modesti per singola
generazione;
c) purtroppo le tecniche di selezione utilizzate, se vanno bene per organismi
più semplici come i moscerini sono del tutto inapplicabili per l’uomo.
Risultati simili sono stati ottenuti con esemplari longevi ottenuti da reincroci con un
vermiciattolo chiamato Caenorhabditis elegans.
Nel 1988 Johnson riportò che la mutazione di un solo gene, chiamato age-1 può far
aumentare del 70% la vita media di C. Elegans.
Probabilmente la proteina prodotta dal gene non mutato inibisce la produzione di
agenti antiossidanti.
2
Il gene dell’invecchiamento HGPS
La progeria di Hutchinson-Gilford(HPGS) è una rarissima patologia che provoca alle
sue giovani vittime un invecchiamento precoce, da 5 a 10 volte maggiore rispetto al
normale decorso.
Questa malattia porta i bambini a morte a un ‘età media di 13 anni. La mutazione si
verifica nel gene LMNA, contenente le informazioni per la costruzione di due
proteine, le lamine A e C.
Gli studi delle sequenze del DNA dei geni LMNA nei pazienti affetti dalla progeria
(HPGS), hanno permesso ai ricercatori di individuare un’anomalia in un segmento
chiamato esone 11, che consiste in una sostituzione di una base di DNA.
Danni al DNA come causa dell’invecchiamento
Il danno al genoma è un problema universale per la vita. Il materiale genetico, DNA,
è instabile in condizioni fisiologiche.
Il DNA è inoltre continuamente assalito da sostanze chimiche reattive. Quando
intervengono danni al DNA esistono due possibilità:
• i danni possono essere riparati
• i danni possono rimanere all’interno del genoma della cellula
L’accumulo di danni al DNA è associato con un declino nell’espressione dei geni
oltre che un declino nelle funzioni cellulari, dei tessuti e degli organi → ciò
implica una relazione di causa ed effetto tra danni al DNA e declino funzionale che
viene definito INVECCHIAMENTO.
L’accumulo di mutazioni somatiche durante tutta la vita non gioca il ruolo
determinante nell’iniziare il processo di invecchiamento nei mammiferi ma
l’invecchiamento è una conseguenza non adattativa del danno al DNA. Quando
i geni che codificano per enzimi deputati ad inattivare gli agenti che danneggiano il
DNA o che riparano il danno non funzionano più, subentra l’invecchiamento.
3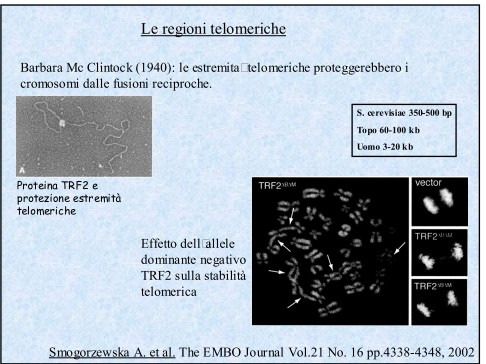
Ma quanti geni ci sono esattamente?
Il dibattito è caldo:
• Human genome consortium ha identificato 30'000
• Celera 30'000
• HGC x Celera -> 45'000 (only 15'000 overlap)
• Over 150'00 unique EST cDNAs have been identified
Quanti prodotti di geni sono mancanti?
4
5
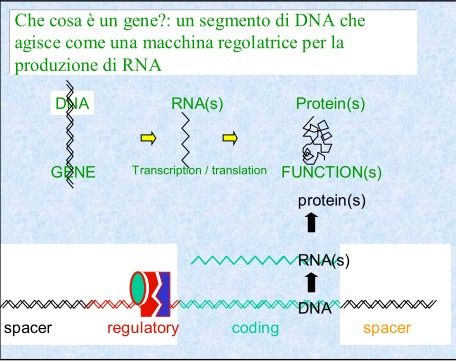
Ogni volta che le cellule si dividono devono duplicare i propri geni in modo
accurato, affinchè ogni cellula figlia riceva un corredo genico completo.
Nel 1972 un ricercatore notò che le DNA polimerasi non erano in grado
di copiare il filamento di DNA per tutta la sua lunghezza fino all’estremità.
È qui che interviene l’attività della telomerasi.
6
Durante gli anni settanta uno studioso sovietico associò la cessazione
programmata della divisione cellulare al problema della duplicazione terminale,
proponendo che le cellule somatiche umane potrebbero non correggere
l’accorciamento cromosomico che ha luogo durante la duplicazione del DNA.
La divisione cessa quando le cellule si accorgono che i loro cromosomi sono
diventati troppo corti?
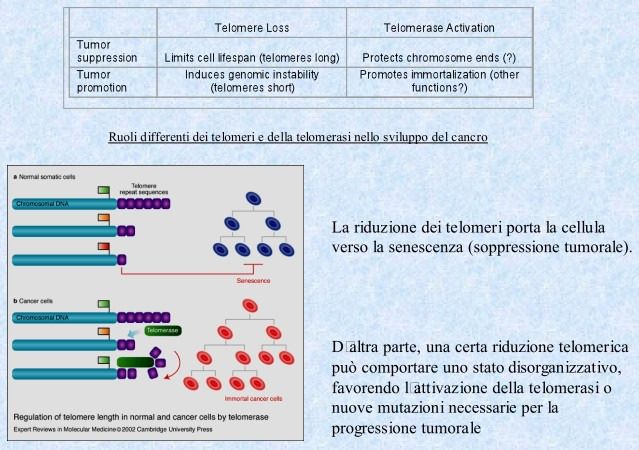
Telomerasi e cancerogenesi…
• Natura e attività della telomerasi;
• L’attività telomerasica e l’insorgenza/progressione dei tumori;
• Proteine accessorie e loro contributo nell’insorgenza tumorale;
• Approccio alla diagnosi tumorale basata sull’attività telomerasica;
• Telomerasi e terapie antitumorali.
7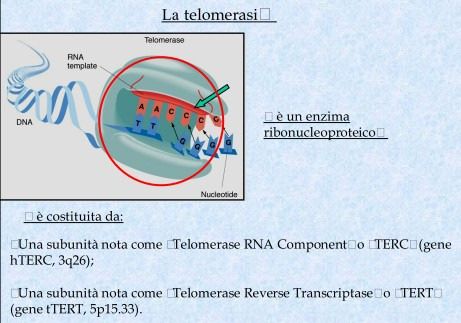
Modello sperimentale della telomerasi
La telomerasi viene prodotta normalmente dalle cellule della linea germinale
dell’embrione in via di sviluppo.
Tuttavia una volta che l’organismo si è completamente formato, la telomerasi
viene soppressa in molte cellule somatiche e i telomeri si accorciano allorchè
queste cellule si riproducono.
Quando la lunghezza dei telomeri raggiunge un valore soglia, viene emesso un
segnale che impedisce alle cellule di dividersi ulteriormente.
Se invece le mutazioni genetiche che promuovono il cancro bloccano l’emissione
di questi segnali o consentono alle cellule di ignorarli, queste ultime non
entrano nella normale fase di senescenza e continuano a dividersi.
Infatti se le cellule non perdono del tutto i telomeri, questi ridimensionati in
lunghezza possono essere salvati, facendo acquisire alla cellula l’immortalità
caratteristica delle cellule tumorali.
8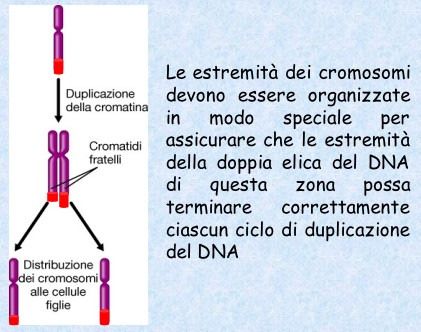
Secondo il modello della regolazione della lunghezza dei telomeri nell’organismo
umano, le cellule della linea germinale producono telomerasi con cui antengono
costante la lunghezza dei telomeri.
Per contro molte cellule somatiche sono prive di telomerasi e quindi con il passare
del tempo i loro telomeri si accorciano → la maggior parte delle cellule somatiche
diventa senescente e cessano di dividersi.
E le cellule aberranti?
Attività telomerasica e terapia dei tumori
• Inibitori della trascrittasi inversa (modelli virali);
• Inibizione della processività della telomerasi;
• Blocco dell’accesso della telomerasi al telomero tramite G-rich oligonucleotidi
• (effetto sui telomeri delle cellule normali???);
• Mutanti hTERC;
• Telomerasi come antigene per l’immunoterapia
9
Espressione specifica di costrutti con hTERT-geni terapeutici solo nelle cellule
con attività telomerasica (espressione tipica delle cellule neoplastiche);
Geni virali e distruzione cellule→
Studio della telomerasi e della sua attività
• Fisiologia cellulare (controllo ciclo cellulare);
• Coinvolgimento telomerasi nell’insorgenza tumorale;
• Terapia antitumorale (modelli murini vie diverse di controllo della replicazione
cellulare, lunghezza telomerica e attività telomerasica!);
• Studio patologie associate a disfunzione telomerasica;
• Sfruttamento attività telomerasica per sintesi molecolari su larga scala.
APOPTOSI
La biologia della morte cellulare e le relazioni con l’invecchiamento
L'omeostasi tra la produzione di nuove cellule e l' eliminazione delle cellule
"vecchie" è uno degli obiettivi principali di ogni organismo pluricellulare. Inoltre
è comunemente accettato che in tali organismi, l' eliminazione di alcune cellule sia
un prerequisito essenziale per il differenziamento in linee cellulari con diversa
specializzazione; tale fenomeno ha un carattere di importanza fondamentale, ad
esempio, nel corso dello sviluppo fetale.
La disregolazione dell’apoptosi è stata implicata in malattie autoimmuni, nella
sindrome da immunodeficienza acquisita e in altre infezioni virali e batteriche,
così come in patologie neurodegenerative, cardiopatie e cancro.
Una disregolazione dell’apoptosi potrebbe contribuire anche al processo di
invecchiamento..
La maggior parte, se non tutte le cellule hanno la capacità di auto-distruggersi
mediante l'attivazione di un programma di suicidio al quale sembrano fare
eccezione solo cellule particolarmente specializzate quali quelle del muscolo
cardiaco e del tessuto nervoso. Questo programma suicida prende il nome di
apoptosi o morte cellulare programmata.
Un aiuto alla comprensione di come differenti segnali possano avere un effetto
cooperativo nell' attivazione di un comune programma di morte cellulare, è venuto
dagli studi di genetica sul controllo dell'apoptosi nel moscerino della frutta,
Drosophila melanogaster.
10Alcuni ricercatori sono giunti all'isolamento di un gene chiamato reaper, che è in
grado di integrare l'informazione proveniente da diversi segnali.
Mutanti per delezione nel gene reaper mostrano soppressione dell’ apoptosi.
Lo studio del gene bcl-2 ha permesso di chiarire le basi molecolari della
insensibilità delle cellule tumorali all'apoptosi. Il gene bcl-2 è coinvolto nella
traslocazione cromosomica osservata nella maggior parte dei linfomi follicolari
umani.
Nel periodo immediatamente successivo alla sua scoperta, questo gene era
ritenuto un oncogene, ma studi più recenti hanno dimostrato che la proteina Bcl-2
ha scarsa o nulla capacità a promuovere la proliferazione delle cellule in
vitro.
Studi sull 'interazione tra la proteina p53 e Bcl Bcl-2 hanno permesso di chiarire i
meccanismi molecolari alla base dell' effetto parodosso di Bcl-2. Infatti, in alcuni
casi l'iperespressione di questa proteina impedisce alle cellule di entrare in
apoptosi.
Morte cellulare ed infezioni virali
Il suicidio di una cellula infettata rappresenta un meccanismo di difesa cellulare per
prevenire la propagazione virale. Le cellule T citotossiche riconoscono ed uccidono
le cellule che presentano peptidi virali associati a molecole MHC (major
histocompatibility complex) di classe I.
Le cellule T citotossiche esercitano il loro effetto inducendo apoptosi mediante
attivazione del recettore Fas sulla superficie della cellula target o mediante
introduzione di proteasi, come granzima B.
Numerosi virus, come adenovirus o papilloma virus hanno tuttavia sviluppato
meccanismi in grado di bloccare l'apoptosi.
Affinchè l'infezione degli adenovirus abbia successo è necessario che il virus
esprima la proteina E1B 19KD. Questa proteina presenta somiglianze con Bcl-2, di
cui è nota la funzione anti-apoptotica.
11Puoi anche leggere

















































