COSTRIZIONI SECONDARIE - Siti fragili comuni - Elearning
←
→
Trascrizione del contenuto della pagina
Se il tuo browser non visualizza correttamente la pagina, ti preghiamo di leggere il contenuto della pagina quaggiù
Siti fragili comuni
- si trovano in tutti gli individui sani analizzati con una frequenza di espressione
variabile da individuo ad individuo;
- rappresentano un fenomeno inerente alla struttura stessa della cromatina.
I siti fragili comuni sono stati scoperti durante lo studio della sindrome dell’X-fragile. Infatti studiando individui sani e
individui affetti, inibendo parzialmente la replicazione del DNA, si osservò la presenza di lesioni non casuali su tutti gli
individui.
I siti fragili comuni rappresentano quindi una caratteristica intrinseca della struttura cromatinica.
COSTRIZIONI SECONDARIE
I siti fragili comuni o costituzionali sono presenti come un carattere costante in tutti gli individui,
seppur con frequenze di espressione diverse. Rappresentano quindi un fenomeno inerente alla
struttura stessa della cromatina.
La percentuale di metafasi che presentano SFC può variare da individuo a individuo e raramente sono
state osservate frequenze di espressione superiori al 30%.
1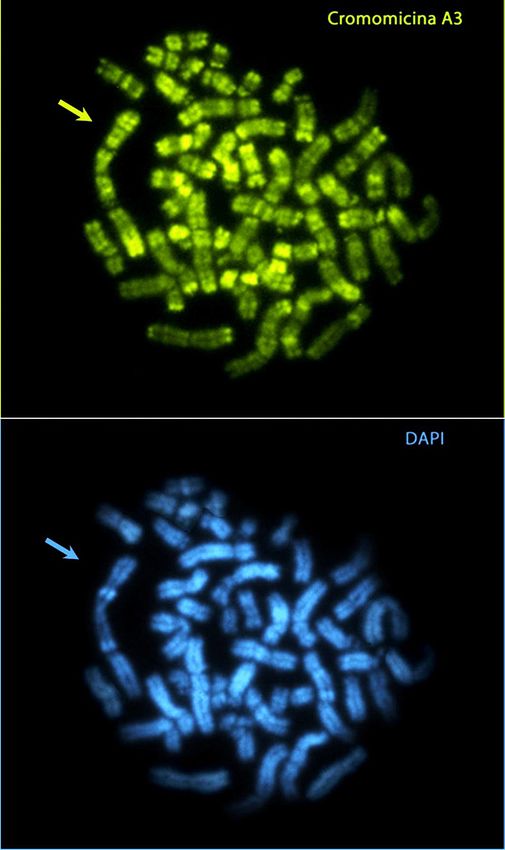
Durkin & Glover, Chromosome Fragile Sites. Annu. Rev. Genet. 2007. 41:169–92. doi:10.1146/annurev.genet.41.042007.165900
Fig. 2 Locations of common fragile sites.
I SF comuni osservati più
frequentemente sono:
FRA3B ( 3p14)
FRA16D (16q23)
FRA7G ( 7q31)
FRA7H ( 7q32)
FRA6F ( 6q21)
FRAXB ( Xp22)
FRA7I ( 7q36)
Xp22.3
Xp211-p21.1
2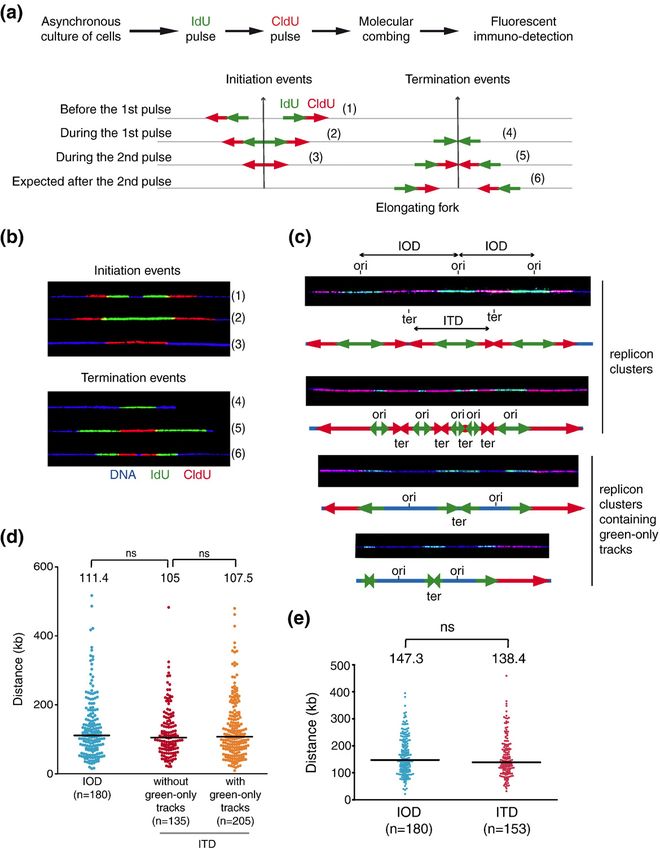
Localizzazione di alcuni Siti Fragili Comuni
in linfociti e fibroblasti
Debatisse et al., Trends in Genetics, 28 (1):22-32, 2012
Note that most fragile
sites differ between
lymphocytes and
fibroblasts.
Exceptions are FRA16D
(16q23.2), a major site in
both cell types; FRA3B
(3p14.2), a major site in
lymphocytes but a minor one
in fibroblasts; and 7q31.1, a
minor site in both cell types.
Chromosomes ideograms are
a schematic representation
of bands visible after Giemsa
staining and were taken
from: http://
www.pathology.washington.ed
u.
The sites with the highest frequencies of breakage
in each cell type are boxed (major sites). 3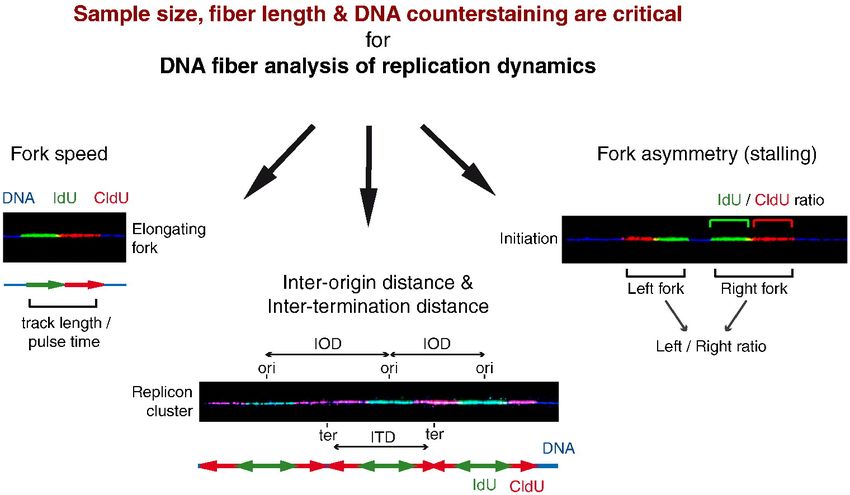
Induzione citogenetica
• Afidicolina inibitore della DNA polimerasi α e δ
• 5-azacitidina analogo della citosina incorporata nel DNA
• BrdU analogo della timidina incorporata nel DNA
SITI FRAGILI COMUNI
(In realtà estese “regioni fragili”)
Analisi delle sequenze di alcuni siti
• presenti in tutti gli individui fragili comuni
indotti da Afidicolina
• componenti “normali” della struttura
cromosomica (costrizioni secondarie)
• “regioni” fragili (fino a 10 Mb)
Assenza di sequenze consenso
• nessun minisatellite associato
FRAGILITA’ ???
Per i siti fragili comuni non è stata trovata alcuna espansione di tri- o dinucleotidi in grado di spiegare la fragilità…
E’ stato quindi usato un approccio che guardasse alla strutturale tridimensionale di queste regioni piuttosto che alla
struttura primaria…
…ed è emerso che queste estese regioni mantengono delle caratteristiche che sono altamente condivise:
• Regioni ricche in AT e particolarmente flessibili
• Ricche in elementi ripetuti
• Colocalizzano con sequenze MAR
• Sono spesso regioni a replicazione tardiva o ritardata
4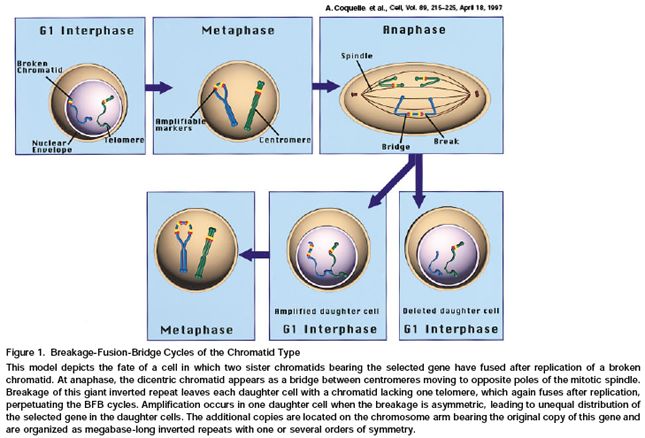
Flessibilità
Regioni ricche in AT
Bend.it – TwistFlex – FlexStab
Misurano la flessibilità come variazione dell’angolo di
twist tra coppie di basi adiacenti della doppia elica
Identificano mediante un algoritmo le sequenze
flessibili (100 bp -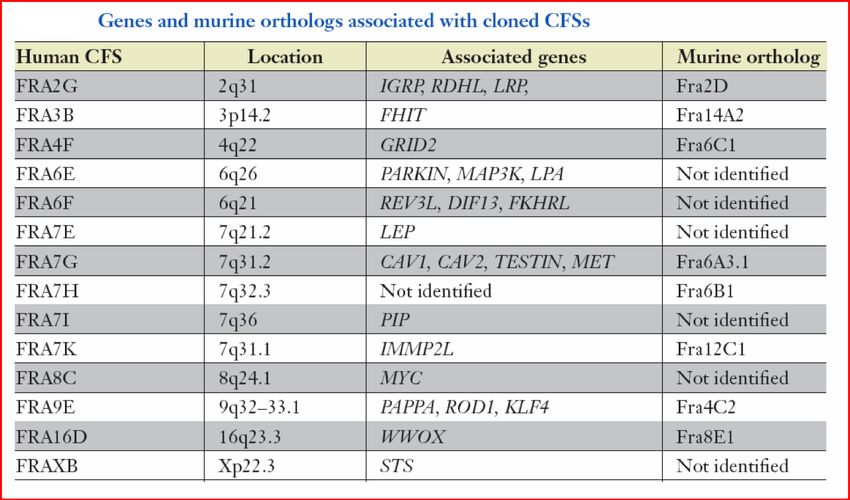
Elementi ripetuti interspersi
Regioni di attacco alla Matrice
MAR - Matrix Attachment Region
Percentuale elevata di elementi LINE, SINE, MIR, MARwiz
MER, LTR e TRASPOSONI
Formazione di strutture intra-molecolari stabili in grado di
- interferire con la replicazione
- produrre delezioni in presenza di meccanismi di riparo
Rallentamento della replicazione del DNA
MAR – Potenziali origini di replicazione
6
Durkin & Glover, Chromosome Fragile Sites. Annu. Rev. Genet. 2007. 41:169–92. doi:10.1146/annurev.genet.41.042007.165900
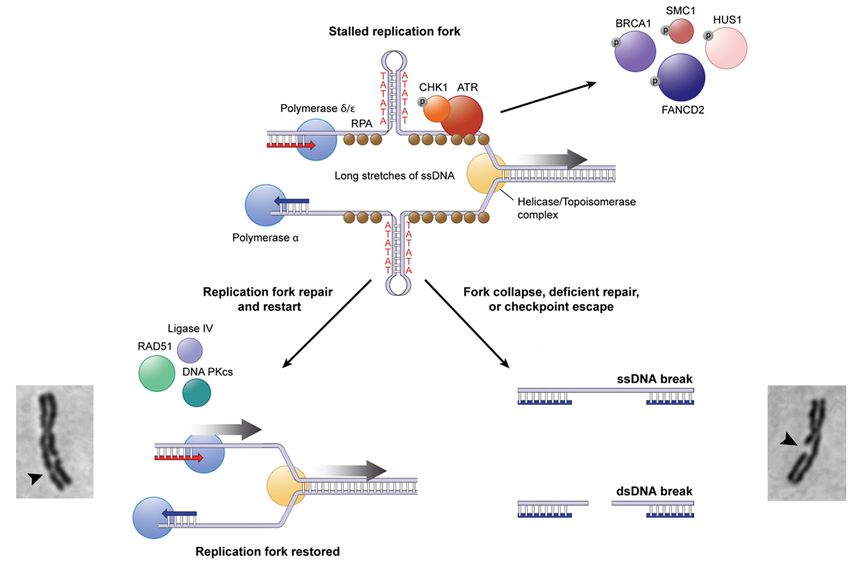
Fig. 3 Modello proposto per l’espressione
dei siti fragili comuni
Una forca replicativa in stallo o in ritardo
(es. trattamento con APH):
l’elicasi continua ad agire sul dsDNA
producendo ssDNA che può formare
strutture secondarie stabili che rallentano
ulteriormente la replicazione del DNA
The ssDNA binding protein RPA (brown) coats the unreplicated ssDNA and recruits the DNA damage
response checkpoint proteins, including ATR (red), which activate S-phase or G2/M checkpoints.
Repair of these regions by proteins including RAD51 and DNA-PK restores replication fork progression.
However, sometimes these regions escape checkpoint activation or are left unrepaired, resulting in an
unreplicated region that can appear as a fragile site on metaphase chromosomes or lead to a DSB.
7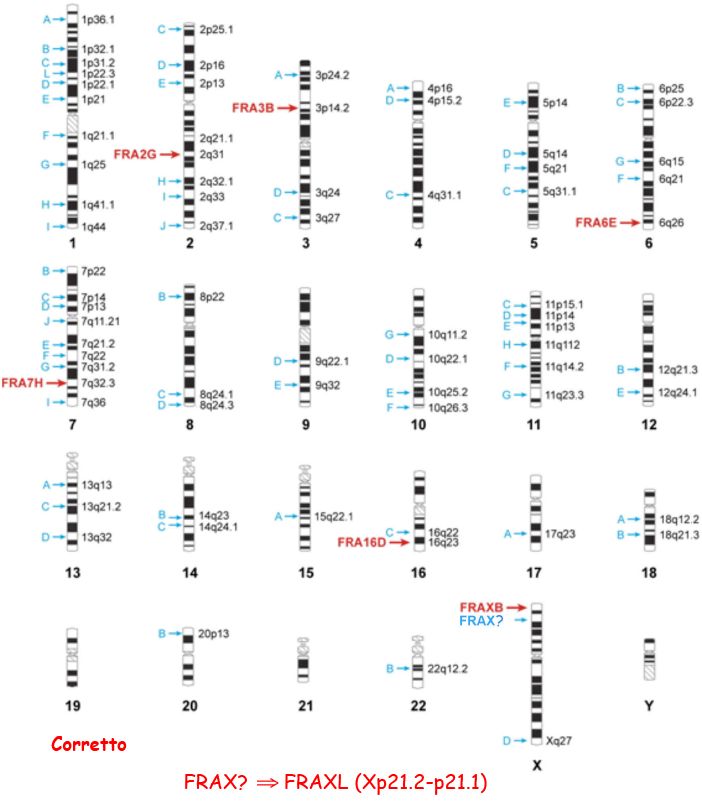
• Il significato biologico ed evolutivo dei SF rimane enigmatico. Sono siti evolutivamente conservati
[dal lievito ai mammiferi] ma potenzialmente dannosi per l’organismo…
• Nell’ultimo decennio è stato ipotizzato un ruolo meccanicistico svolto dai SF nell’insorgenza e nella
progressione tumorale.
• Coincidente localizzazione delle bande in cui sono presenti oncogeni e quelle in cui sono mappati
taluni SF (Sutherland 1985).
• In seguito a trattamento con 16 mutageni e carcinogeni il 67% dei punti di rottura cromosomica
coincide con la localizzazione di SF (Yunis and Soreng, 1987).
I SF sono i siti di inizio e reintegrazione nell’amplificazione genica intracromosomica tramite cicli
di rottura-fusione-ponte in condizioni di ipossia (Coquelle et al. 1997).
Nei primi anni 2000 è stata analizzata la frequenza d’espressione dei siti fragili comuni in pazienti affetti dal cancro al
colon, alla mammella, ai polmoni e al collo e testa. In tutti i casi è stato riscontrato un significativo aumento, presente
anche nei parenti di primo grado non affetti da neoplasia.
Ciò lascia intravedere la prospettiva di approntare dei test per misurare la suscettibilità genetica ad alcune forme
tumorali.
Caratteristiche principali dei siti fragili comuni
• punti caldi per lo scambio tra cromatidi fratelli
• siti di traslocazioni cromosomiche
• siti di delezioni cromosomiche
• siti di amplificazione genica (ciclo rottura-fusione-ponte)
• siti di integrazione per virus e DNA plasmidico
Regioni normalmente stabili in coltura, ma in presenza di agenti inibitori della replicazione, manifestano 8
caratteristiche di DNA altamente instabile e ricombinogeno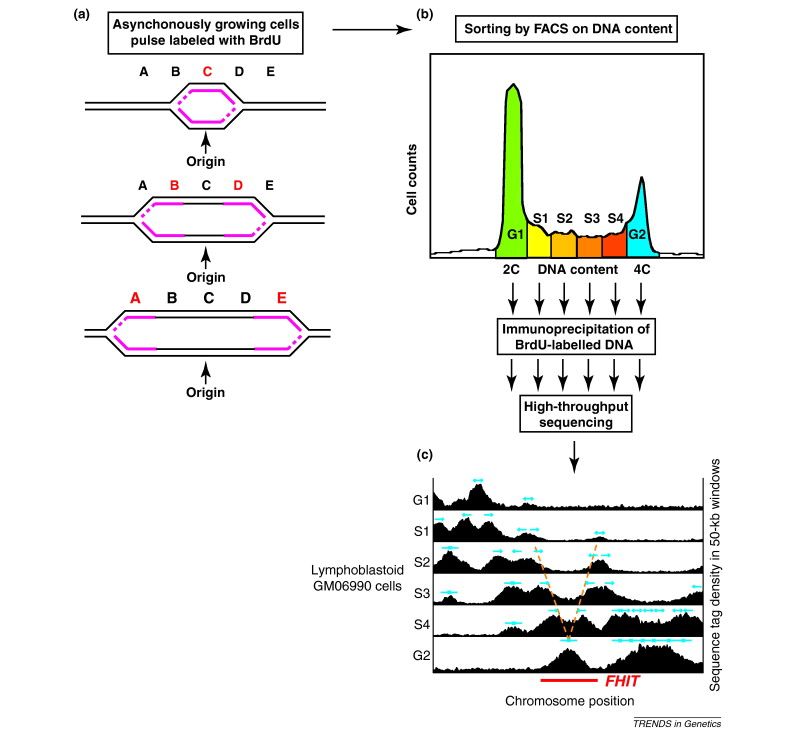
Siti fragili, instabilità genomica e replicazione
Irony-Tur Sinai & Kerem, DNA replication stress drives fragile site instability. Mutat Res Fund Mol Mech Mutagen, 808, March 2018,56-61.
https://doi.org/10.1016/j.mrfmmm.2017.10.002
Highlights
• DNA replication stress drives genome instability and initiates tumorigenicity
• Nucleotide deficiency and deregulated origin firing lead to replication stress.
• CFSs exhibit intrinsic sensitivity to replication stress conditions.
• Under stress the replication along CFSs is delayed and even uncompleted.
• CFSs are preferentially unstable in pre-cancerous lesions and during cancer development.
In this review article, we focus on the early events initiating DNA replication stress and the
preferential sensitivity of common fragile sites (CFSs) to this stress.
CFSs are specific genomic regions within the normal chromosomal structure, which appear as gaps
and breaks in the metaphase chromosomes of cells grown under mild replication stress conditions.
The main characteristics predisposing CFSs to instability include late replication timing, delayed
replication completion, failure to activate additional origins, origin paucity along large genomic
regions, collision between replication and transcription complexes along large genes, and the
presence of AT-dinucleotide rich sequences.
The contribution of these features to instability at CFSs during early cancer development is
discussed.
9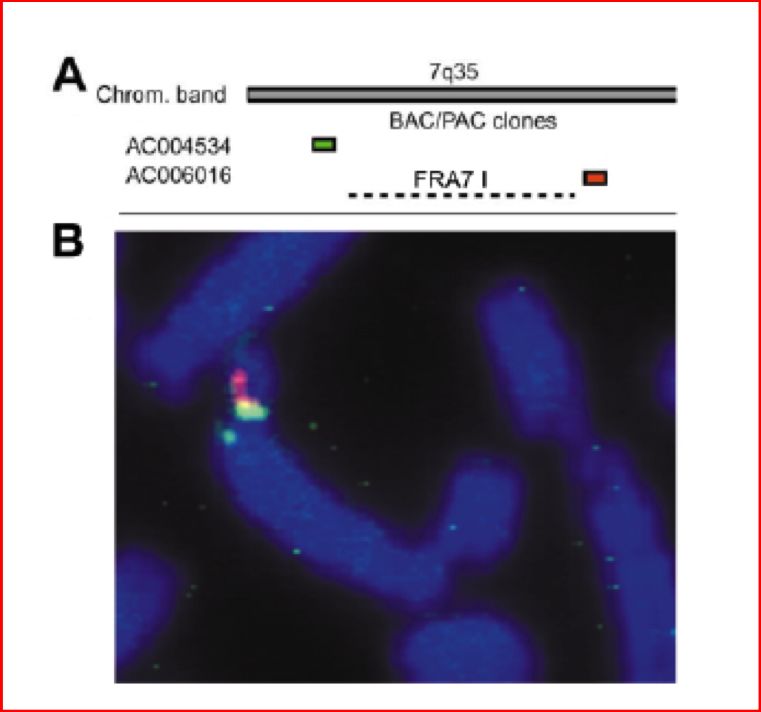
Irony-Tur Sinai & Kerem, DNA replication stress drives fragile site instability. Mutat Res Fund Mol Mech Mutagen, 808, March 2018,56-61.
https://doi.org/10.1016/j.mrfmmm.2017.10.002
Fig 1. Schematic illustration summarizing the early events initiating DNA
replication stress and the main features predisposing CFSs to instability.
10Méchali M.: Eukaryotic DNA replication origins: many choices for appropriate answers. Nature Reviews Molecular Biology, 11:728-738, Oct 2010
Replication fork:
When replication starts, the opened DNA forms two branched structures on both sides of the
replication origin that resemble forks. Fork progression is mediated by the action of DNA
helicases that unwind the DNA and facilitate the movement of the DNA synthesis machinery.
In Escherichia coli, DNA replication starts from a single, sequence-specific element, and the
speed of the two replication forks (60 kb min–1) keeps pace with a rapid cell cycle (less than
30 min).
The human genome is 700 fold larger than the E. coli genome, but the replication fork speed
is 20 fold slower (2–3 kb min–1). Thus, it would take at least 20 days to achieve a single division
if there was one origin per chromosome.
Not all origins are activated at the same time; their activation follows the specific timing
of DNA replication during the cell cycle.
11Méchali M.: Eukaryotic DNA replication origins: many choices for appropriate answers. Nature Reviews Molecular Biology, 11:728-738, Oct 2010
Figure 1 | Replication origins.
a | At each replication origin, DNA synthesis starts
with short RNA primers that are synthesized by
DNA polymerase α.
As DNA synthesis always occurs in the 5′–3′
direction, one strand of the DNA (the leading
strand) will be synthesized continuously, whereas
the other strand (the lagging strand) will be
synthesized discontinuously by short RNA-primed
DNA fragments.
Two other DNA polymerases (δ and ε) are
recruited for the elongation of lagging and
leading strands, respectively.
b | Activation of replication origins during
S phase.
Pre-replication complexes (preRCs) are assembled
at replication origins during G1 phase. Activation
of replication origins occurs throughout S phase,
some during early (1 and 2), and some in mid (3) or
late (4) S phase.
preRC: complesso di preRepliCazione 12Méchali M.: Eukaryotic DNA replication origins: many choices for appropriate answers. Nature Reviews Molecular Biology, 11:728-738, Oct 2010
Figure 3 | Different types of DNA
replication origins.
Potential DNA replication origins are
set during mitosis–G1 phase by the
assembly of pre-replication complex
(preRC) proteins.
The selection of the origins that will
be activated at the next S phase
occurs at G1 phase and may vary
according to the cell fate or
environmental conditions.
Four examples of DNA replication origin positions are shown in different cells in a growing cell population.
A cluster of flexible origins contains origins that can be used differently in different cells. Their use
could increase or decrease according to physiological or abnormal growth conditions.
Inactive or dormant origins are rarely used or are not used at all.
Constitutive origins are fixed origins that are always set at the same position by chromatin
or transcriptional constraints.
Replication stress can activate dormant origins or increase the use of flexible origins,
resulting in an increased number of origins per replication cluster.
13Méchali M.: Eukaryotic DNA replication origins: many choices for appropriate answers. Nature Reviews Molecular Biology, 11:728-738, Oct 2010
Figure 4 | Checkpoint
control of replication
origins firing.
During DNA replication,
a DNA lesion caused by
stresses such as ultraviolet
damage, oxidative damage,
genotoxic drugs or growth
medium deprivation may
slown down or even
arrest progression of the
replication fork.
Replication stress can induce or increase a signal transduction cascade, called the checkpoint response,
which tries to maintain the integrity of the replication forks and facilitate DNA repair in coordination
with the cell cycle.
The checkpoint signalling through Ataxia Telangiectasia mutated-Related (ATR) results in the inhibition
of late origins.
The frequency of origin usage in clusters of early origins might also be negatively regulated by t
he ATR pathway, through a lateral inhibition of activated origins on the potential neighbouring origins.
However, replication stress might induce a second pathway that cancels this inhibitory effect
on early origins, resulting in activation of dormant early origins.
14Técher et al., Replication Dynamics: Biases and Robustness of DNA Fiber Analysis. Journal of Molecular Biology null 2013.
http://dx.doi.org/10.1016/j.jmb.2013.03.04
Fig 1. Principles of replication dynamics analyses.
(a) Scheme of the protocol used for double pulse labeling of replication forks in
asynchronous cell cultures. Successive pulses with IdU and CldU enable recognition
of ongoing forks, initiation and termination events. Green and red arrows represent
neo-synthesized DNA, respectively, labeled with IdU and CldU.
The labeling pattern depends on whether the considered event occurs:
- before the first pulse,
- during the first pulse,
- during the second pulse or
- after the second pulse.
Six different patterns of initiations and terminations are expected, numbered 1–6. 15Técher et al., Replication Dynamics: Biases and Robustness of DNA Fiber Analysis. Journal of Molecular Biology null 2013.
http://dx.doi.org/10.1016/j.jmb.2013.03.04
► Fork speed is subjected to 10–15% of experimental variability.
► Sample size impact on fork speed and IOD measurement.
► Fiber length impact on measured fork speed, IOD and fork asymmetry.
► Thirty percent of fibers are broken at the level of replication signals.
► DNA counterstaining is required to assess replication parameters.
IdU: iododeoxyuridine ITD: inter-termination distance
CldU: chlorodeoxyuridine IOD: inter-origin distance
16Debatisse et al., Common fragile sites: mechanisms of instability revisited. Trends in Genetics, 28(1):22-32. 2012.
http://dx.doi.org/10.1016/j.tig.2011.10.003
Common fragile sites (CFSs) are large chromosomal regions prone to breakage
upon replication stress that are considered a driving force of oncogenesis.
Recent studies show that delayed completion of DNA replication depends on a
regional paucity in initiation events.
Because the distribution and the timing of these events are cell type dependent,
different chromosomal regions can be committed to fragility in different
cell types.
These new data reveal the epigenetic nature of CFSs and open the way to a
reevaluation of the role played by these sites in the formation of chromosome
rearrangements found in tumors from different tissues.Debatisse et al., Common fragile sites: mechanisms of instability revisited. Trends in Genetics, 28(1):22-32. 2012.
http://dx.doi.org/10.1016/j.tig.2011.10.003
Fig I Analysis of replication in the fragile histidine triad ( FHIT ) gene region by DNA combing technique.
DNA molecules several hundred kb long are
stretched on a solid support and are all
oriented in the same direction.
Fig. Ia, newly synthesized DNA is successively
pulse-labeled in vivo for the same length of time
with two thymidine analogs, IdU then CldU.
Fig Ib, cells are immediately recovered and
embedded in agarose plugs.
DNA is gently purified in the plugs then released
in solution upon agarose melting.
Combing is performed on silanized coverslips that
are dipped into the DNA solution. DNA molecules
bind spontaneously to the coverslip surface,
preferentially by their extremities. When the cover
slip is pulled out of the solution, the meniscus exerts
a constant force on the anchored molecules when
they come out at the surface.
IdU= iododeoxyuridine; CldU= chlorodeoxyuridinw Parallel DNA fibers are aligned over the cover slip
with a constant stretching factor (1 mm = 2 kb).
Fig Ic, tracks of DNA containing IdU or CldU are revealed in two different colors using specific fluorescent
antibodies. The length of each track reflects the speed of the replication fork in the corresponding region
because the labeling time is constant. In addition, the double labeling permits determination of fork direction.
Given that replication is initiated bidirectionally from each origin, initiation events are revealed by two divergent
forks. Termination events occur by the merging of two convergent forks. To study specific loci, fluorescence in
situ hybridization is performed with a series of probes revealed in a third color, allowing unambiguous alignment
of the molecules relative to the map of the locus. In the case of the FHIT gene illustrated here, 29 probes
(green tracks) grouped in five distinguishable motifs were necessary to cover this 1.5 Mb-long gene.Debatisse et al., Common fragile sites: mechanisms of instability revisited. Trends in Genetics, 28(1):22-32. 2012.
http://dx.doi.org/10.1016/j.tig.2011.10.003
Fig 1. Schematic representation of replication dynamics in the
core region of FRA3B in human lymphocytes and fibroblasts.
The core region is delimitated by orange lines and cell-cycle phases are indicated on the left with S phase subdivided in four
fractions (Box 2). The core is poor in initiation events in lymphocytes and during an unperturbed S phase, it is replicated by
long-traveling forks emanating from origins located in the flanking regions that fire around mid-S. Convergent forks merge at
the end of S phase or in G2 phase, resulting in late completion of the core replication. Under conditions of replication stress,
long-traveling forks are impeded and the replication of the core is incomplete when the cells enter mitosis, despite the
eventual occasional firing of some inefficient origins within the core.
By contrast, in fibroblasts, many initiation events occur within the core during the second half of unperturbed S phase and
are responsible for late completion of replication in this region. Under replication stress conditions, the decrease in fork
speed is compensated by an increase in the density of initiation events, resulting in replication completion before the cells
enter mitosis.Debatisse et al., Common fragile sites: mechanisms of instability revisited. Trends in Genetics, 28(1):22-32. 2012.
http://dx.doi.org/10.1016/j.tig.2011.10.003
Box 2.
The Repli-Seq technique
Box 2.
The Repli-Seq technique
Figure I.
Replication timing in the
fragile histidine triad
(FHIT) gene region revealed
by the Repli-Seq technique.Debatisse et al., Common fragile sites: mechanisms of instability revisited. Trends in Genetics, 28(1):22-32. 2012.
http://dx.doi.org/10.1016/j.tig.2011.10.003
Localizzazione di alcuni Siti Fragili Comuni
in linfociti e fibroblasti
Note that most fragile
sites differ between
lymphocytes and
fibroblasts.
Exceptions are FRA16D
(16q23.2), a major site in both
cell types; FRA3B (3p14.2), a
major site in lymphocytes but a
minor one in fibroblasts; and
7q31.1, a minor site in both cell
types.
Chromosomes ideograms are a
schematic representation of
bands visible after Giemsa
staining and were taken from:
http://
www.pathology.washington.edu.
The sites with the highest frequencies of breakage
in each cell type are boxed (major sites).Il cancro come malattia genetica
Mutazioni a carico di due principali classi di geni:
Geni soppressori tumorali – ostacolano la crescita del tumore, la loro perdita
favorisce la progressione tumorale (mutaz. in omozigosi)
Proto-oncogeni – geni cellulari la cui alterazione qualitativa o quantitativa favorisce
la tumorigenesi
22Ruolo meccanicistico dei SF nella trasformazione tumorale
1. Siti preferenziali di integrazione di virus oncogeni
Alterazione di proto-oncogeni cellulari
Interruzione di geni soppressori tumorali
2. Delezione di geni soppressori tumorali
es. FRA3B e il gene FHIT
FRA3B
• regione 3p14.2
• sito fragile indotto dall’ afidicolina. E’ quello che ha la maggiore frequenza di espressione nel genoma
umano,
• si estende per circa 500 Kb
• sito d’integrazione del papillomavirus-16 umano (HPV16)
Questa regione è deleta in omozigosi in diverse forme tumorali, compresa una traslocazione reciproca con il cromosoma 8
presente nel carcinoma renale familiare (hRCC).
Coincidenza con un sito già caratterizzato dall’integrazione di HPV-16 nel carcinoma della cervice uterina.
Tutto ciò suggerisce la presenza di un gene tumor-suppressor
23Nella regione FRA3B è stato identificato il gene FHIT (Fragile Histidine Triad)
Il locus FHIT ( 1.5 Mb) è composto da 10 esoni, l’esone 5 si trova nella regione fragile deleta in omozigosi in molte neoplasie.
L’ mRNA risulta assente o ridotto in molti tessuti tumorali.
La proteina FHIT (16.8 kDa) è un enzima con attività di diadenosina trifosfato idrolasi che scinde ApppA in ADP e ATP in
vitro.
Studi su interazioni proteina-proteina, linee cellulari
tumorali e topi knockout suggeriscono un ruolo
della proteina FHIT nella proliferazione cellulare e
nell’ apoptosi.
Sono state sequenziate 870 kb del locus FHIT/FRA3B, dall’esone 3 all’esone 7 del gene.
Il locus è ricco in AT e ha poche sequenze Alu. Non sono presenti altre sequenze geniche.
Sono presenti regioni di alta flessibilità, considerate responsabili della fragilità cromosomica.
24FRA16D (16q32.2)
Le rotture e le decondensazioni si estendono in una regione di almeno 1 Mb.
E’ tra i più grandi SF clonati fino a questo momento.
I marcatori presenti in FRA16D risultano deleti in tumori della prostata, della mammella e ovarici indicando che la
perdita di questa regione può essere importante nello sviluppo o nella progressione di queste neoplasie.
Inoltre una traslocazione t(14q32;16q23) è stata osservata in più del 25% di tutti i mielomi multipli.
Nella regione FRA16D è stato identificato il protooncogene MAF, la sua espressione risulta aumentata in seguito alla
traslocazione presente in alcuni mielomi multipli.
Tre laboratori hanno identificato in modo indipendentemente il gene FOR (WWOX o WOX1) nella regione FRA16D.
La proteina codificata è un’ossidoriduttasi in grado di prendere parte al processo di apoptosi e di interagire con p53
e altre proteine coinvolte nella tumorigenesi.
FRA16D (16q23.2) - WWOX
Regione deleta in diversi tipi di tumori
Delezioni mappano all’interno del gene WWOX
WWOX è stato classificato come gene soppressore tumorale probabilmente coinvolto nei processi apoptotici
Topi KO per WWOX sviluppano tumori precocemente rispetto a topi di controllo
Sono in corso studi sulla funzione di questo gene come onco-soppressore, si conosce la sua funzione come 25
ossidoreduttasi, ma non si sono ancora trovati interattori certi.O'Keefe & Richards. Common chromosomal fragile sites and cancer: Focus on FRA16D. Cancer Letters, 232(1):37–47, 2006.
http://dx.doi.org/10.1016/j.canlet.2005.07.041
Fig. 1. FRA16D associated DNA instability in cancer and the WWOX (FOR) gene.
Diagram represents the cytogenetic
location of FRA16D and the corresponding
locations of Loss-Of-Heterozygosity (LOH)
regions mapped in prostate and breast
cancers; translocation breakpoints in
multiple myeloma and homozygous deletions
in adenocarcinomas.
The location of the WWOX (FOR) gene
including its 780 Kb intron is also depicted.
26FRA7G (7q31.1) Le rotture indotte dall’afidicolina si verificano in una regione di almeno 300 kb.E’ presente in essa una sequenza retrovirale endogena. Coincidente localizzazione con regioni delete in varie forme tumorali quali il carcinoma del colon e l’adenocarcinoma ovarico. Presenza di un putativo gene soppressore tumorale. Nella regione sono presenti i geni codificanti la caveolina 1 e la caveolina 2, Il primo gene è in grado di indurre apoptosi e sopprimere la crescita in alcune linee tumorali ma non risulta alterato nelle linee tumorali che presentano la delezione della regione FRA7G. E’ stato di recente identificato un gene che presenta omologia di sequenza con il gene murino della testina. Il gene è espresso in tutti i tessuti umani e risulta mancante o in quantità ridotta in alcune neoplasie analizzate. Resta ancora da analizzare il ruolo effettivamente svolto dal gene nella tumorigenesi. L’analisi di sequenza ha rivelato regioni ad alta flessibilità e bassa stabilità. FRA7H (7q32.3)) E’ stata identificata e sequenziata una regione di 161 kb.Sono state così riscontrati numerosi tratti di DNA in grado di assumere strutture inusuali e sequenze con alta flessibilità e bassa stabilità. FRA6F( 6q21) Si estende per circa 1200 Kb, con due punti principali di rotture cromosomiche. La sequenza è ricca di elementi ripetuti e regioni ad alta flessibilità. Presenta delezioni in varie forme di leucemia e tumori solidi (Parkin gene). FRAXB (Xp22.3) Localizzato in Xp22.3. Si estende su una regione genomica di almeno 500 kb in cui mappano tre geni conosciuti, uno dei quali codifica per la steroide sulfatasi microsomiale Sono state riscontrate delezioni a carico della regione fragile in vari tumori primari e linee cellulari tumorali FRA7I (7q36) Presente sul cromosoma 7 in posizione q35, è stato caratterizzato da poco tramite FISH e sembra estendersi per più di 2 Mb. A monte del SF si trova il gene PIP (prolactin-inducible protein) sopra-espresso nell’80% dei tumori alla mammella primari e metastatici. FRA9E (9q32) Si estende per ben 9 Mb.E’ localizzato in 9q32.33.1. Nella stessa regione è situato il gene PAPPA (pregnancy-associated plasma protein-A) 27 Questo gene presenta LOH nel 50% dei tumori ovarici
Geni presenti all’interno di siti fragili comuni coinvolti in processi tumorali
28Ciullo et al., Initiation of the breakage–fusion-bridge mechanism through common fragile site activation in human breast cancer cells: the model of PIP
gene duplication from a break at FRA7I. Human Molecular Genetics, 2002, Vol. 11(23):2887-2894.
3. Amplificazione di proto-oncogeni
Amplificazione selettiva di una regione intracromosomica
tra due siti fragili adiacenti mediata dal ciclo di
rottura-fusione-ponte
FRA7I (7q36) e il gene PIP
⇒
⇒
FRA7I
29Coquelle et al., Cell, Vol. 89, 215–225, April 18, 1997
Fig 1. Breakage-Fusion-Bridge Cycles of the Chromatid Type
This model depicts the fate of a cell in which two sister chromatids bearing
the selected gene have fused after replication of a broken chromatid.
At anaphase, the dicentric
chromatid appears as
a bridge between
centromeres moving to
opposite poles of the
mitotic spindle.
Breakage of this giant inverted repeat leaves each daughter cell with a chromatid lacking one telomere,
which again fuses after replication, perpetuating the BFB cycles. Amplification occurs in one daughter
cell when the breakage is asymmetric, leading to unequal distribution of the selected gene in the
daughter cells. The additional copies are located on the chromosome arm bearing the original copy of
30
this gene and are organized as megabase-long inverted repeats with one or several orders of symmetry.Ciullo et al., Initiation of the breakage–fusion-bridge mechanism through common fragile site activation in human breast cancer cells: the model of PIP
gene duplication from a break at FRA7I. Human Molecular Genetics, 2002, Vol. 11(23):2887-2894.
Model for the generation of the inverted
duplication found in T47D cells
Duplicazione di PIPPelliccia et al: A molecular cytogenetic analysis: KG-1a leukemia cell line. Oncology Letters 4:237-240, 2012. DOI:10.3892/ol.2012.709
Cell line AML KG-1aRP11-79E5
Chr 1 Der(1) RP11-438F14
RP11-385F5
RP11-79E5
RP11-79E5: 1q12 (h_sat) RP11-79E5
RP11-385F5: 1q43 (≈10Mb dal Tel)
RP11-438F14: 1q44 (≈ 3Mb dal Tel)
RP11-385F5
RP11-438F14
RP11-385F5
RP11-438F14RP11-79E5: 1q12 (h_sat) RP11-296O14: 1q24.2 RP11-434B7: 1q32.3
Quindi…
• I siti fragili comuni sono estese regioni di fragilità che arrivano a coprire anche diverse
Mb del genoma
ü Rappresentano una caratteristica strutturale del cariotipo [costrizioni secondarie] e
la loro conservazione fa supporre un loro ruolo biologico importante
Ø Non si conosce la causa della fragilità in queste regioni, ma è stato dato un ruolo
meccanicistico nello sviluppo tumorale
Referenze
- Durkin SG, Glover TW 2007. Chromosome fragile sites. Annu Rev Genet. 2007;41:169-92.
- Glover TW et al 2005. Mechanisms of common fragile site instability. Hum Molec Genetics, 14
Spec No.2:R197-R205. 2005 Oct 15. Review.
- Schwartz M et al. 2006. The molecular basis of common and rare fragile sites.
Cancer Letters 231:13-26. Review
- Debatisse M et al. 2012. Common fragile sites: mechanisms of instability revisited.
Trends in Genetics, 28 (1):22-32, 2012.
- Ozeri-Galai et al. 2012. The complex basis underlying common fragile site instability in cancer
Trends in Genetics, 28 (6):295-302
- Irony-Tur Sinai & Kerem, 2018. DNA replication stress drives fragile site instability.
Mutat Res Fund Mol Mech Mutagen, 808, 56-61, March 2018.
35Puoi anche leggere

















































