Valutazione dello stimolo circadiano in presenza di differenti filtri selettivi
←
→
Trascrizione del contenuto della pagina
Se il tuo browser non visualizza correttamente la pagina, ti preghiamo di leggere il contenuto della pagina quaggiù
Università degli Studi di Napoli “Federico II” Scuola Politecnica e delle Scienze di Base Area Didattica di Scienze Matematiche Fisiche e Naturali Dipartimento di Fisica “Ettore Pancini” Laurea triennale in Ottica e Optometria Valutazione dello stimolo circadiano in presenza di differenti filtri selettivi Relatori: Candidato: Prof. Laura Bellia Laura Fiore Prof. Antonio Sasso Matricola M44/363 A.A. 2018/2019 1
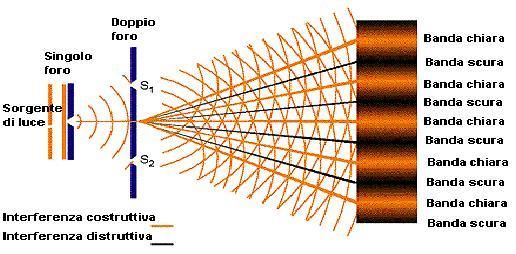
Indice Introduzione……………………………………………………………………………...5 Capitolo 1: Effetti visivi e non visivi della luce…………………………………………6 1.1 Il sistema visivo……………………………………………………………...6 1.1.1 Anatomia del bulbo oculare………………………………………..6 1.1.1.1 La retina.……………………………………………………..10 1.1.1.1.1 Fotorecettori: coni e bastoncelli………………………12 1.2 Sensibilità spettrale del sistema visivo……………………………………..14 1.3 Il sistema non visivo o circadiano…………………………………………..16 1.3.1 Sensibilità spettrale del sistema circadiano……………………….16 Capitolo 2: La scoperta di un nuovo fotorecettore: ipRGC ……………………………20 2.1 La risposta delle ipRGC alla luce…………………………………………..20 2.1.1 Sensibilità spettrale delle ipRGC…………………………………22 2.1.2 Il riflesso pupillare alla luce………………………………23 2.2 I ritmi circadiani…………………………………………………………….25 2.3.1 Il funzionamento dell’orologio biologico………………………..25 2.3.2 Alterazioni dell’orologio circadiano nell’uomo…………………27 Capitolo 3: Metodologia di misurazione e presentazione delle misure………………...29 3.1 Misurazione della trasmittanza dei filtri ottici……………………………...29 3.1.1 Spettrofotometro………………………………………………….29 3.1.1.1 Schema ottico di Lamba 25, 35, 45……………………..30 3.1.1.1.1 Reticolo di diffrazione………………………...32 3.1.1.1.1.1 Diffrazione…………………………..33 3.1.1.1.1.2 Dispersione………………………….34 3.1.2 Filtri ottici………………………………………………………...36 3.1.2.1 Trasmittanza e Assorbanza……………………………...37 3.2 Metodologia di acquisizione dei poteri di distribuzione spettrale delle radiazioni e loro stimolo circadiano…………………………………………….40 3.2.1 Luce circadiana…………………………………………………...40 3.2.2 Presentazione e descrizione delle radiazioni selezionate…………42 2
3.3 Valutazione dello stimolo circadiano in presenza di filtri selettivi in ambienti interno ed esterno……………………………………………………………….45 3.3.1 Rappresentazione delle radiazioni filtrate e presentazione dei valori ottenuti………………………………………………………………….46 3.3.2 Valori ottenuti dal filtraggio del simulatore della luce naturale per differenti illuminamenti………………………………………………...53 Capitolo 4: L’illuminazione degli ambienti per il benessere dell’individuo 4.1 Approccio tradizionale ai progetti d’illuminazione 4.2 Nuovo ruolo dei sistemi di illuminazione negli ambienti 4.3 Approccio teoretico per introdurre nuove proprietà per la caratterizzazione dell’ambiente luminoso 4.4 Definizione e misurazione di quantità radiometriche circadiane Conclusioni Bibliografia Ringraziamenti 3

“Dolce è la luce e agli occhi piace vedere il sole”, Ecclesiaste (11,7). 4

Introduzione La luce è un elemento essenziale per la vita sulla Terra; essa è responsabile della maggior parte dei processi chimici e biologici che governano sia il mondo vegetale che animale. Recenti scoperte hanno ampliato il campo degli effetti che questo elemento ha sull’essere umano e sugli altri viventi del pianeta: un nuovo fotorecettore osservato all’interno della retina, ha mostrato una dipendenza tra radiazione elettromagnetica e ritmi fisiologici, regolati dall’orologio circadiano, che appare essere indissolubile. La comunità scientifica introduce dunque il concetto di luce circadiana, il quale non si riferisce più a quella luce che ci permette di osservare gli oggetti a noi circostanti, ma una luce che è responsabile degli effetti non-visivi, attraverso il suo stimolo circadiano. Tale lavoro di tesi ha come obiettivo la valutazione di quest’ultimo, in presenza di differenti filtri ottici selettivi, di diverse sorgenti artificiali che caratterizzano l’illuminazione moderna degli ambienti interni e della radiazione che illumina gli ambienti esterni, la luce del giorno. La progettazione dell’illuminazione ha da sempre l’obiettivo di rendere ottimali le prestazioni visive, avendo cura esclusivamente degli aspetti legati al comfort visivo. Con l’avvento delle scoperte legate al nuovo ruolo della luce, concernenti le implicazioni che essa ha sulla salute e sul benessere, si stanno ricercando progettazioni finalizzate non più soltanto a rendere ottimale la prestazione visiva richiesta ma improntate anche a una più completa e corretta illuminazione naturale e artificiale degli ambienti. Si sta assistendo cioè al passaggio da un’impostazione più strettamente tecnica a una integrata, tuttavia soltanto quando sarà raggiunta una totale consapevolezza e conoscenza dell’impatto che la luce ha sugli aspetti biologici, verrà garantita una corretta progettazione illuminotecnica. Molte sono ancora le questioni irrisolte; quel che appare certo però è che nel futuro un’illuminazione salubre e sostenibile dovrà prevedere una progettazione orientata sia agli aspetti visivi (performance e comfort) che a quelli circadiani, integrata dagli aspetti di efficienza energetica e sostenibilità ambientale. Al fine di comprendere suddetti concetti che attorniano lo scopo di tale lavoro, l’argomentazione di tale tesi è stata suddivisa in quattro capitoli: nel primo capitolo sono introdotti gli effetti visivi e non-visivi della luce rispettivamente sul sistema visivo e non visivo umano, caratterizzati ognuno da una distinta sensibilità spettrale. Nel secondo capitolo invece, sono approfonditi i concetti concernenti il sistema non- visivo: dall’elemento fondamentale per la sua attivazione, l’ipRGC, al ritmo circadiano. Nel terzo capitolo, sono presentate le misure ottenute e spiegata la metodologia di misurazione. Infine, nel quarto capitolo è introdotto l’approccio tradizionale ai progetti di illuminazione, basato sull’applicazione delle grandezze fotometriche, e la metodologia per integrare in esso le grandezze circadiane, in modo da ottenere un’illuminazione salubre sotto tutti gli aspetti. 5
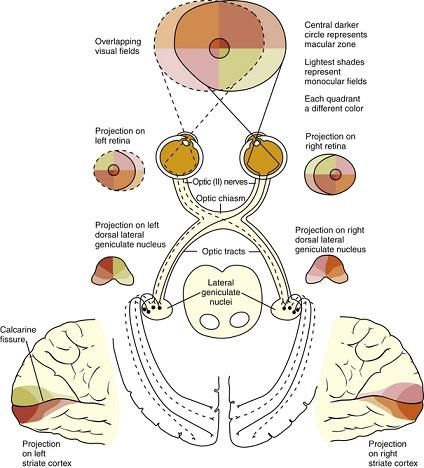
CAPITOLO 1 Effetti visivi e non visivi della luce Il dizionario di fisica “The Penguin” (1977) definisce la luce come: “L’agente che causa una sensazione visiva quando essa cade sulla retina dell’occhio”. La specie umana è considerata “visiva”, poiché percepiamo ed esploriamo l’ambiente esterno attraverso gli occhi, e con essi trasmettiamo all’ambiente stesso sensazioni, pensieri, e vari elementi della comunicazione non verbale [1]. Tra i cinque sensi, infatti, quello della vista è per l’uomo il più importante. Infatti è su ciò che vediamo, più che sulle altre percezioni, che si basa gran parte delle nostre azioni coscienti, ed inoltre l’estensione delle aree celebrali dedicate alla visione è molto più ampia di quelle dedicate agli altri sensi [2]. Gli esseri umani recepiscono la luce attraverso gli occhi, all’interno dei quali i due fotorecettori, coni e bastoncelli, permettono di catturarne le informazioni visive. Catturando tali informazioni, i fotorecettori attivano il sistema visivo attraverso il quale l’uomo è capace di valutare l’ambiente circostante, attraverso la percezione di spazio e dettagli [3]. Recenti scoperte, avvenute negli ultimi trent’anni, hanno dimostrato che l’illuminazione della retina influenza anche molti aspetti della fisiologia e del comportamento dell’uomo. Tale dipendenza ha origine nell’occhio ma non fa parte degli aspetti della visione, in quanto indipendente da specifiche configurazioni spaziali dello stimolo luminoso, ed infatti può sopravvivere anche in soggetti non vedenti [4]. Insomma, possediamo una sorta di “visione cieca” che non genera immagini, e che tuttavia è in grado di inviare segnali precisi al cervello. Nell’uomo tale visione è affidata alle speciali cellule gangliari retiniche [5]. 1.1 Il sistema visivo Il sistema visivo (Figura 1) è l’insieme delle strutture coinvolte nella “conversione della luce nella sensazione e nelle percezioni della visione” [6]. Nell’essere umano esso si compone di: • Bulbo oculare ed annessi; • Nervo ottico; • Tratto ottico; • Pretetto, Ganglio ciliare, Nucleo oculomotore accessorio; • Collicolo Superiore; • Corpo genicolato laterale; • Radiazioni ottiche o calcarine; • Corteccia visiva, suddivisa in primaria, secondaria e terziaria; • Corteccia associativa. 6
FIGURA 1: COMPONENTI DELL’APPARATO VISIVO UMANO, DALL’OCCHIO ALLE AREE DI PROIEZIONE DELL’IMMAGINE DEL CERVELLO [7] . Il principale organo del sistema visivo è l’occhio, il cui compito è di acquisire informazioni sull’ambiente circostante attraverso la luce. L’occhio dell’essere umano raccoglie la luce proveniente dall’ambiente, regolandone l’intensità attraverso un diaframma (l’iride), e, mediante un sistema regolabile di lenti, la focalizza per formare un’immagine sulla retina. Tale immagine verrà successivamente trasformata in segnali elettrici che, tramite il nervo ottico, sono diretti al cervello per l’elaborazione e l’interpretazione [8]. Nel paragrafo seguente, verrà riportata la struttura anatomica del bulbo oculare attraverso l’analisi delle sue tre componenti principali. 1.1.1 Anatomia del bulbo oculare Il bulbo oculare occupa la porzione anteriore della cavità orbitaria. Esso ha approssimativamente la forma di una sfera schiacciata leggermente dall’alto e dal basso, con un diametro trasversale di 23,6 mm ed un diametro verticale di 23,2 mm. E’ possibile distinguere due poli diametralmente opposti, uno anteriore corrispondente al centro della superficie corneale ed uno posteriore sulla superficie posteriore della sclera, nell’intervallo tra l’emergenza del nervo ottico. I due poli sono congiunti da un asse immaginario chiamato asse anatomico che coincide con l’asse ottico (linea ideale, che passando per il centro ottico raggiunge la fovea centrale). L’asse anatomico non coincide però con la linea visiva la quale rappresenta la linea che unisce la fovea centrale (zona dotata di maggior sensibilità morfoscopica) ed il punto nodale dell’occhio (centro ottico dell’occhio posto dinanzi alla superficie posteriore del cristallino) [9]. La struttura dell’occhio può essere suddivisa in tre componenti principali: 7
1. Tre tonache (Figura 2), che costituiscono la parete del bulbo oculare: • La tonaca fibrosa che è lo strato più esterno, è a sua volta divisa in due parti: la sclera e la cornea. La sclera consiste di un tessuto connettivo collagene perforato da vasi sanguigni e nervi che ricopre la maggior parte della superficie oculare; La cornea invece, è la porzione anteriore trasparente della sclera modificata che consente l’ingresso della luce all’interno dell’occhio. • La tonaca vascolare costituisce invece lo strato intermedio, viene definita anche uvea per la somiglianza ad un acino d’uva ed è costituita da tre parti: la coroide, il corpo ciliare e l’iride. La coroide è uno strato riccamente vascolarizzato e pigmentato, posto dietro la retina. Anteriormente si ispessisce per diventare corpo ciliare, FIGURA 2: SEZIONI DELLE TRE TONACHE CHE che sostiene il cristallino e forma un RIVESTONO IL BULBO OCULARE [10]. anello muscolare intorno ad esso, il muscolo ciliare che ne controlla la tensione, modulata per garantire la messa a fuoco degli oggetti; Il corpo ciliare sostiene anche l’iride, un diaframma regolabile che controlla il diametro della pupilla, la sua apertura centrale. • La tonaca interna infine è formata dalla retina, che riveste internamente i due terzi posteriori del bulbo oculare [11]. 2. Un sistema diottrico (Figura 3) costituito da elementi trasparenti di forma peculiare e caratteristiche differenti, che permettono l’ingresso della luce, in modo da creare un’immagine reale, capovolta e con i lati invertiti, a fuoco sul piano retinico, di un oggetto compreso in un campo visivo di oltre 200°. Le sue caratteristiche complessive e il suo potere variano continuamente a causa dell’accomodazione, tuttavia le strutture FIGURA 3: ELEMENTI DEL SISTEMA che lo compongono gli conferiscono DIOTTRICO DELL’OCCHIO [12]. complessivamente un potere positivo di 60 diottrie circa [13]. Tali strutture sono: • Il film lacrimale. E’ una struttura liquida formata dalle lacrime prodotte dall’apparato lacrimale, che ricopre come una pellicola la cornea e la congiuntiva palpebrale bulbare. Oltre ad avere una funzione refrattiva, se pur minima, essendo una superficie con indice di rifrazione proprio, possiede 8
una funzione ottica: migliorare la trasparenza della superfice corneale, mediante il suo strato mucoso. • La cornea, precedentemente citata, rappresenta la lente più potente dell’apparato visivo (43 D). Ha il compito di garantire il passaggio dei raggi luminosi verso le strutture interne facendole convergere verso la fovea, ma anche di proteggere le sub-strutture da abrasioni e agenti patogeni. • L’umor acqueo, un fluido sieroso prodotto dal corpo ciliare che occupa due spazi, la camera posteriore compresa tra l’iride e il cristallino, e la camera anteriore posta tra cornea e iride. • Il cristallino, una lente dalla forma biconvessa che permette di mettere a fuoco gli oggetti prossimali. E’ sostenuto dietro la pupilla da un anello di fibre definito legamento sospensore, che lo ancora al corpo ciliare. Ha un potere che con l’accomodazione varia dalle 16 D alle 20 D. • Il corpo vitreo, infine, è un gel trasparente che invade uno spazio cospicuo chiamato camera vitrea, posta alle spalle del cristallino. Esso sostiene il bulbo e mantiene la retina uniformemente compressa contro la superficie interna della cavità; essenziale per mettere a fuoco le immagini sulla retina. 3. Le componenti nervose (Figura 4), che comprendono il nervo ottico e la retina. • Il nervo ottico è il secondo nervo di 12 paia di nervi cranici. E’ costituito da fibre nervose che originano dalle cellule ganglionari retiniche e, convergendo verso la papilla ottica, danno luogo alla formazione del tronco nervoso. Lascia l’orbita attraverso il canale ottico, raggiungendo il chiasma, in cui si assiste ad una parziale decussazione delle fibre nervose. La maggior parte degli assoni del nervo ottico giunge al corpo genicolato laterale, da dove le informazioni visive vengono FIGURA 4: COMPONENTI NERVOSE DELL’OCCHIO [14]. trasmesse alla corteccia visiva. • La retina è una membrana sottile trasparente adesa solo in due punti: il disco ottico, ed il suo margine anteriore dentellato, l’ora serrata. Il resto della retina è tenuto contro la parte posteriore del bulbo oculare in modo uniforme, dalla pressione del corpo vitreo [11]. 9
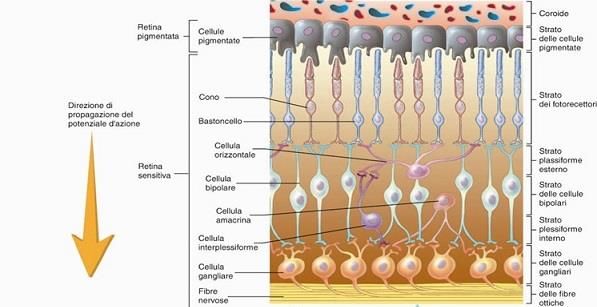
1.1.1.1 La retina La retina è paragonabile ad un sensore di una macchina fotografica digitale: ha il compito di catturare l’informazione luminosa e di trasformare le immagini in impulsi nervosi che il nervo ottico trasmette poi al cervello. E’ considerata come un estroflessione del diencefalo; essa è un fine tessuto nervoso sensibile alla luce che forma tutto il rivestimento interno del bulbo oculare (Figura 5), dal punto di entrata del nervo ottico al margine pupillare dell’iride e viene FIGURA 5: RETINA E LE SUE DUE AREE suddivisa in tre sezioni: una parte ottica, una NOTEVOLI, LA FOVEA E LA MACULA [15]. ciliare (che aderisce al corpo ciliare) e la parte iridea. Le ultime due sezioni, costituite esclusivamente da tessuto epiteliale, formano la parte cieca della retina; solo la parte ottica, quindi, possiede i fotorecettori, un particolare tipo di cellule nervose in grado di rilevare la presenza di luce; a tale livello ha origine la sensazione visiva. Curiosamente, la retina appare come capovolta, infatti le radiazioni devono attraversare l’intero spessore prima di raggiungere i fotorecettori. Per questo motivo la retina, nonostante le numerose cellule che la compongono, è trasparente. Una volta che i fotorecettori sono stimolati, il segnale che da essi origina percorre la rete dei neuroni retinici lungo il nervo ottico, per raggiungere il cervello. Struttura Santiago-Ramon y Cajal, attraverso le sue ricerche, ha permesso di individuare precisamente la struttura della retina. Questa si presenta divisa in dieci livelli principali (Figura 6): • Strato epiteliale pigmentato, costituito da cellule a base esagonale e propaggini ciliate verso l’interno; • Strato dei fotorecettori, contengono il segmento esterno dei fotorecettori; • Membrana limitante esterna, costituita dalle cellule gliali di Muller; • Strato nucleare esterno, comprendono il segmento interno dei fotorecettori e i loro nuclei; • Strato plessiforme esterno, sito delle sinapsi tra fotorecettori, cellule bipolari ed amacrine; • Strato nucleare interno, formato dai corpi cellulari delle cellule gangliari; • Strato plessiforme interno, luogo delle sinapsi tra cellule bipolari, gangliari e amacrine; • Strato delle cellule gangliari, costituito dai corpi cellulari delle cellule gangliari; • Strato delle fibre ottiche, formato dagli assoni delle cellule gangliari; • Membrana limitante esterna, composta dalle stesse cellule della membrana limitante interna, racchiude strutture nervose e vasi. 10
FIGURA 6: STRATI RETINICI [16]. Generalità morfologiche La retina è una struttura complessa, formata dalle connessioni di cinque tipi principali di cellule, e rappresenta l’unica parte del sistema nervoso centrale che consente l’ispezione visiva. Essa è composta da uno strato di epitelio pigmentato e una parte nervosa: il primo aderente alla lamina fusca della coroide, la seconda posta più internamente al bulbo. 1. L’epitelio pigmentato è un tessuto formato da un gruppo di cellule aventi una struttura di base esagonale e delle propaggini ciliate verso l’interno che entrano in contatto con le porzioni più esterne dei fotorecettori. Tale strato ha due funzioni fondamentali: una funzione ottica resa possibile dalle cellule che lo compongono in quanto, essendo ricche di pigmento, rendono opaca la cavità posteriore oculare; e una funzione metabolica essendo sede di notevoli scambi con i fotorecettori 2. La porzione nervosa della retina consta di tre strati principali di cellule. Partendo posteriormente all’occhio, questi sono composti da: • Cellule gangliari. Sono i neuroni più massivi della retina, monostratificati in prossimità del corpo vitreo. Rappresentano i neuroni di secondo ordine della via visiva essendo l’ultimo livello dell’elaborazione retinica. Esse sono connesse alle cellule amacrine e bipolari e i loro assoni sono connessi con il corpo genicolato, dopo aver formato il nervo ottico. Morfologicamente non vi sono grandi differenze tra le cellule gangliari, ma ve ne sono notevoli nella funzionalità; non contribuiscono alle immagini visive ma rivelano solo l’intensità luminosa attraverso il loro pigmento sensitivo chiamato melanopsina; • Cellule bipolari. Sono neuroni di primo ordine che contraggono sinapsi con i fotorecettori retinici; infatti il segnale originatosi nei coni e bastoncelli prosegue grazie a tali cellule. 11
A tale livello agisce un primo controllo, che attua il primo perfezionamento della codificazione dei fotorecettori, infatti il segnale di questi ultimi può essere modulato o completamente inibito nel passaggio alle cellule bipolari; • Fotorecettori. Vi sono due cellule recettoriali differenti: coni e bastoncelli, che hanno la funzione di convertire l’assorbimento di onde elettromagnetiche in un segnale chimico, e quindi elettrico, attraverso il processo di fototrasduzione. Verranno approfonditi nel seguente paragrafo. Vi sono altre cellule retiniche ma esse non formano degli strati propri: • Le cellule orizzontali e amacrine che formano connessioni orizzontali tra bastoncelli, coni e cellule bipolari; giocano ruoli diversi nell’aumentare la percezione del contrasto, dei margini degli oggetti e dei cambiamenti di intensità della luce. • Le cellule gliali e astrociti che compongono una parte cospicua della massa della retina [11,13]. 1.1.1.1 Fotorecettori: coni e bastoncelli I fotorecettori (o fotocettori) sono cellule nervose specializzate, sensibili alla luce. La loro fotosensibilità è resa possibile dall’espressione dei pigmenti in essi contenuti che, per effetto delle radiazioni ottiche, subiscono trasformazioni biochimiche, dalle quali ha origine l’impulso nervoso, che successivamente viene smistato ai centri della visione localizzati nel sistema nervoso centrale. Come suddetto, nella retina distinguiamo due tipi di cellule fotorecettoriali: coni e bastoncelli (Figura 7) localizzate nello strato più esterno della retina, a contatto con l’epitelio pigmentato. Coni e bastoncelli differiscono sia dal punto di vista anatomico che, in maggior misura, funzionale. Entrambe le loro strutture possono essere suddivise in due segmenti, separati da una ristretta zona contenente nove paia di microtubuli: uno esterno che contiene il pigmento fotosensibile, e uno interno che costituisce l’apparato metabolico. Alla base di quest’ultimo si origina poi il corpo cellulare, che contiene il nucleo, e processi che istaurano sinapsi con le cellule FIGURA 7: RAPPRESENTAZIONE CONI E retiniche dello strato sottostante. BASTONCELLI [17]. • Bastoncelli. Se ne contano circa 125 milioni, concentrati principalmente nella periferia retinica. Sono responsabili della visione notturna (scotopica), producendo immagini solo in gradazione di grigio, e sono i fotorecettori maggiormente sensibili sia al movimento che alla luce. 12
Vengono così definiti per la forma cilindrica del loro segmento esterno; quest’ultimo è costituito da una pila ordinata di 1000 dischi di vescicole membranose, contenenti il fotopigmento, avvolta da una membrana plasmatica che mantiene i pigmenti in una posizione che assicura FIGURA 8: SEZIONI DEI SEGMENTI ESTERNI DEI CONI E l’assorbimento più efficace BASTONCELLI [18]. della luce (Figura 8). La sostanza fotolabile espressa dai bastoncelli è la rodopsina, fotopigmento avente un picco di sensibilità a circa 507 nm, nell’occhio umano. • Coni. Nella retina umana vi sono circa 6 milioni di coni che si concentrano nella zona centrale della retina (fovea centralis). Sono deputati alla visione diurna (fotopica) e dunque alla percezione dei colori. Anche questi devono il loro nome al segmento esterno che si restringe fino a un punto, definendo una forma a cono. Tale segmento non possiede delle vescicole a se stanti come nei bastoncelli, ma è costituito da un’unica membrana ripiegata su se stessa diverse volte (Figura 8). Il fotopigmento contenuto nei coni è la iodopsina, che si differenzia in tre tipi di ospine spettralmente differenti: iodopsina eritrolabile o eritropsina, iodopsina clorolabile o cloropsina e iodopsina cianolabile o cianopsina. Si distinguono così tre tipi di coni: i coni-S esprimono la cianopsina, pigmento sensibile alle corte lunghezze d’onda, e sono dunque maggiormente sensibili a lunghezze d’onda di circa 420 nm; i coni-M invece hanno un picco di sensibilità a circa 535 nm esprimendo la cloropsina, sensibile alle medie lunghezze d’onda; i coni-L, infine, contengono la eritropsina che ha una sensibilità spostata verso le lunghezze d’onda più lunghe, e hanno un picco di sensibilità a 565 nm [11,13]. “La sensibilità spettrale dell’occhio adattato alla luce dipende dai tre fotopigmenti contenuti nei coni. Poiché la sensibilità assoluta del cono a lunghezza d’onda corta (cono-S) è solo un decimo di quella degli altri due, la sensibilità spettrale della visione fotopica è dominata dagli altri due coni a lunghezza d’onda più lunga. I coni, presi singolarmente, non sono capaci di codificare la lunghezza d’onda di uno stimolo luminoso. Se un cono risponde a un fotone, genera la stessa risposta, indipendentemente dalla lunghezza d’onda di quel fotone; ogni tipo di pigmento di cono può assorbire una vasta gamma di lunghezze d’onda. Il pigmento è più probabile che assorba i fotoni quando la loro lunghezza d’onda è al picco di assorbimento, ma la luce che colpisce il cono all’estremo della sua gamma di assorbimento può ancora generare un’ampia risposta se l’intensità della luce è sufficientemente elevata. 13
Tale proprietà di univarianza di risposta è la ragione per cui la visione in un occhio con un solo pigmento funzionante può essere unicamente monocromatica: con un singolo sistema di pigmenti la distinzione tra i diversi colori e tra le diverse intensità è confusa. Due diversi coni, ciascuno con una diversa ma sovrapposta gamma di sensibilità di lunghezze d’onda, rimuove gran parte delle ambiguità nel codificare la lunghezza d’onda degli stimoli luminosi. Con tre pigmenti sovrapposti (ciò che avviene negli esseri umani), la luce di una singola lunghezza d’onda stimola ciascuno dei tre coni in gradi diversi e la luce di qualsiasi altra lunghezza d’onda stimola questi coni secondo uno schema decisamente diverso.” (Fisiologia medica - Walter F. Boron, Emile L. Boulpaep) 1.2 Sensibilità spettrale del sistema visivo L’occhio non è ugualmente sensibile a tutte le lunghezze d’onda dello spettro visibile, difatti la curva della sensibilità spettrale visiva è approssimativamente a forma di campana, la cui forma e posizione dipendono dai livelli di illuminazione. Sono identificate due forme estreme della curva della sensibilità spettrale; una per livelli di illuminazione moderati e alti, e una per i livelli bassi. Per livelli moderati e alti di illuminazione, la visione è dominata dai coni, vediamo il colore e la sensibilità spettrale è riferita a quella della sensibilità fotopica. L’intervallo dei livelli intermedi di illuminazione tra tali due estremi è chiamato intervallo mesopico, sia coni e bastoncelli lavorano [19]. FIGURA 9: CURVE DELL’EFFICIENZA SPETTRALE DELL’OCCHIO UMANO IN VISIONE FOTOPICA E SCOTOPICA [20]. Visione fotopica Se un fascio di energia elettromagnetica ha un flusso spettrale radiante denotato con FR(λ), l’ammontare del potere radiante o flusso FR nel fascio è dato dall’integrale: ∞ = ∫ ( ) 0 14
dove FR(λ) ha l’unità di watt per unità di lunghezza d’onda. Per la visione fotopica viene utilizzata, invece, tale formula: ∞ = ∫ ( ) ( ) 0 dove la costante Km è conosciuta come la massima efficacia spettrale luminosa della radiazione per la visione fotopica e ha un valore 683,002 lm/W (CIE, 1983). V(λ) è conosciuta come la funzione dell’efficienza spettrale luminosa per la visione fotopica. Essa è stata determinata dalla risposta media di molti soggetti attraverso un piccolo numero di studi, è stata definita dalla CIE nel 1924. V(λ) ha un massimo valore a 555 nm. La funzione V(λ) presenta valori troppo bassi a circa 450 nm a causa dei limiti negli studi dai quali essa è stata sviluppata, attraverso la sua correzione sono considerati essere superati dall’inconvenienza pratica. Dovrebbe essere considerato che le sensibilità individuali relative fotopiche differiscono per un numero di ragioni, incluse le seguenti: 1. L’efficienza luminosa è l’effetto combinato dei tre tipi di coni, in modo che mentre la sensibilità spettrale dei coni può essere la stessa per ogni individuo, il numero relativo dei coni può variare. Oltretutto, i soggetti che hanno difetti nella visione dei colori hanno la mancanza di un tipo di cono (discromatismo) o un tipo di cono il quale pigmento ha un’alterata sensibilità spettrale (anomalia tricromatica); 2. Variazioni nella trasmittanza spettrale attraverso i mezzi oculari e variazioni nella densità di un pigmento giallo nella macula della retina. In particolare, con l’età, la nostra sensibilità all’azzurro decresce perché le nostre lenti assorbono più azzurro e vi sono cambiamenti neurali associati ai coni sensibili alle corte lunghezze d’onda. (Werner, 1990) [20]. Visione mesopica Come decresce il livello di luce dal livello fotopico attraverso quello scotopico, vi è un cambiamento nella sensibilità spettrale relativa che accompagna la transizione della visione dai coni ai bastoncelli. Tale regione di transizione è chiamata regione mesopica e nella sensibilità spettrale relativa è chiamata “Purkinje shift”. Visione scotopica La CIE ha definito la funzione dell’efficienza spettrale luminosa per la visione scotopica V’(λ) nel 1951. Tale funzione ha un massimo valore a 507 nm. Esso è mostrato come con V(λ), in Figura 9. Se noi convertiamo l’energia radiante in luce usando V’(λ), utilizziamo l’equazione (1) con V’(λ) rimpiazzando V(λ), e K’m, la massima efficacia spettrale luminosa della radiazione per la visione scotopica, rimpiazzando Km. 15
Il valore di K’m è 1700,6 lm/W (CIE, 1983), il quale è derivato dalla definizione che 1 lumen fotopico = 1 lumen scotopico, per una sorgente monocromatica con una frequenza di 540×1012 Hz. Il rapporto dei due valori di Km è il rapporto dei valori di V(λ) e V’(λ), alla lunghezza d’onda di 555,016 nm in aria con un indice di rifrazione di 1,00028. Dato che vi è solo una classe di cellule (i bastoncelli) operanti a livelli di luce scotopici, noi ci dovremmo aspettare meno variabilità tra individui per la sensibilità relativa scotopica, rispetto alla sensibilità relativa fotopica [20]. Limiti fotopici, mesopici e scotopici Non vi sono divisioni nette tra le bande dei tre intervalli, il più basso limite di luminanza della visione fotopica è approssimativamente 3 cd/m2, con visione mesopica che si estende da tale livello a 0,03 cd/m2, dopo il quale inizia la visione scotopica [20]. 1.3 Sistema non-visivo o circadiano Recenti studi hanno portato alla scoperta, all’interno dell’occhio, di un’ulteriore tipo di fotorecettore: le cosi dette ipRGCs, cellule ganglionari intrinsecamente fotosensibili. Su tali cellule si basa il funzionamento del sistema non-visivo. Le ipRGCs rispondono lentamente allo stimolo luminoso, sono dotate di ampissimi campi recettivi e sono presenti, nella retina, in numero molto ridotto. A causa delle loro peculiarità, esse non sono in grado di garantire delle funzioni di creazione delle immagini “visive”, differenziandosi così dai coni e bastoncelli, ma grazie alla costanza nella risposta garantiscono la comprensione dei livelli di illuminazione giornaliera dell’ambiente circostante. Difatti, le ipRGCs intervengono nella mediazione di svariati processi di non formazione dell’immagine, riflettendola a livello subconscio [21]. Per suddetti motivi, la risposta delle ipRGCs alla luce viene definita come risposta non- visiva o di non formazione dell’immagine; tale risposta comprende un’ampia varietà di reazioni, di cui la più influente è sicuramente quella di regolazione dell’orologio endogeno circadiano. Dato che la ritmicità circadiana è una caratteristica di praticamente tutti i sistemi psicologici, metabolici e comportamentali, tale fenomeno porta a una vasta gamma di processi biologici, sotto il controllo indiretto della retina. Aldilà di questo, il termine “risposta non-visiva” è usato per abbracciare una crescente lista di effetti in risposta alla luce, che insieme assicurano uno stato fisiologico che sia sincronizzato con il naturale ciclo giorno-notte [22]. 1.4 Sensibilità spettrale del sistema circadiano Nell'essere umano le sensibilità spettrale, spaziale, temporale ed assoluta del sistema circadiano sono molto diverse da quelle del sistema visivo; infatti, dal punto di vista qualitativo, il sistema circadiano ha una soglia più alta per la stimolazione, mentre il sistema visivo ha un picco di sensibilità alle corte lunghezze d'onda e probabilmente risponde molto più lentamente allo stimolo luminoso. 16
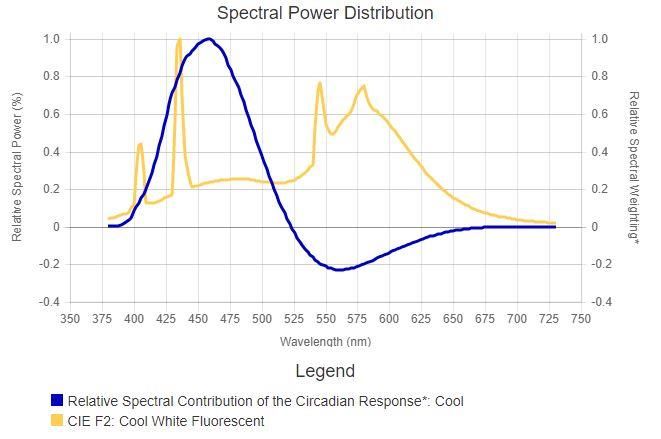
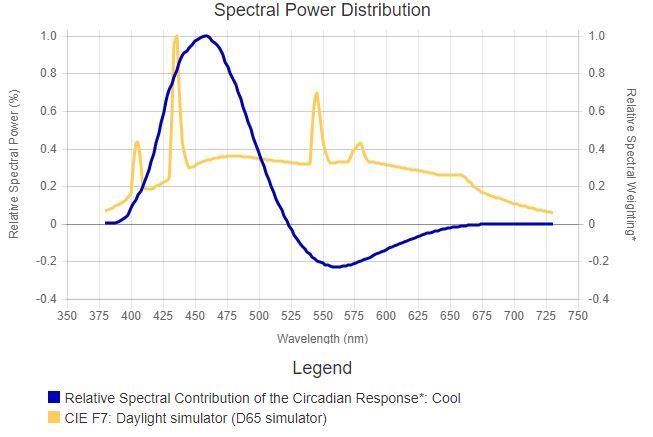
In particolare, vediamo che il sistema circadiano reagisce allo stesso stimolo luminoso in modo opposto a seconda dell’ora del giorno solare in cui avviene la stimolazione: uno stimolo luminoso nel mattino aumenta il tempismo dell'orologio circadiano, di sera invece lo stesso stimolo luminoso lo ritarda. Sebbene possono esser fatte comparazioni qualitative tra i due sistemi, vi è ancora qualche incertezza nell'esatta previsione quantitativa di come il sistema circadiano risponda, attraverso l'esposizione della retina, ai differenti stimoli luminosi. Malgrado tali incertezze, attraverso l’investigazione empirica, è stata convenzionalmente stabilita la sensibilità spettrale del sistema circadiano per lo spettro visibile. In particolare, vari tentativi hanno portato alla costruzione di diversi modelli della sensibilità spettrale che differiscono tra loro sia in termini di complessità, sia per le loro considerazioni circa la neuroanatomia e fisiologia della retina e prima che un modello possa essere rifiutato a favore di un altro sono necessarie verifiche future ed un adeguato livello di controllo sperimentale [4]. 1.4.1 I modelli di Brainard e Thapan I primi due tentativi di costruire un modello della sensibilità spettrale FIGURA 10: DUE SERIE DI STIME DELLA SENSIBILITÀ del sistema circadiano dell’uomo SPETTRALE DEL SISTEMA CIRCADIANO DELL’UOMO BASATA SULLA SOPPRESSIONE DELLA MELATONINA ALLA RISTRETTA provengono dai dati raccolti BANDA DELLO SPETTRO VISIBILE. DUE FUNZIONI SONO indipendentemente da due gruppi di BASATE SU UNA SINGOLA OPSINA, UNA A 460NM E L’ALTRA A ricerca, uno capitanato da Brainard 480NM CIRCA; L’ULTIMA FUNZIONE INVECE, È BASATA SU UN e l'altro da Thapan, entrambi basati ANDAMENTO ARBITRARIO DELLA SENSIBILITÀ SPETTRALE sulla soppressione della melatonina. STIMATA. LE FUNZIONI DELLE SINGOLE OPSINE SONO STATE ENTRAMBE CORRETTE PER TENER CONTO DELLA Sebbene i due gruppi di ricercatori TRASMISSIONE DELLA LENTE SECONDO WYSZECHI E STILES adottano procedure sperimentali [4]. leggermente diverse, entrambi utilizzano un criterio costante per costruire la sensibilità spettrale della soppressione di melatonina causata dalla luce durante le ore notturne. La Tabella 1 e la Figura 10 presentano tali efficienze spettrali, dove lo stimolo luminoso è quantificato in termini di irradianza corneale, come determinato graficamente da Rea et al., usando il 35% come norma di soppressione per le funzioni dose-risposta nelle pubblicazioni originali. TABELLA 1: SENSIBILITÀ RELATIVE A DIFFERENTI PICCHI DI LUNGHEZZA D’ONDA PER LA SOPPRESSIONE DELLA MELATONINA, OTTENUTE DA BRAINARD E THAPAN. SECONDO I DATI DI BRAINARD, TALI EFFICIENZE SPETTRALI SONO DETERMINATE ATTRAVERSO LA DIGITALIZZAZIONE DEI GRAFICI DELLE FUNZIONI DOSE- RISPOSTA PER OGNI LUNGHEZZA D’ONDA DELL’INTERVALLO VISIBILE. E’ STATO STIMATO IL POTERE RADIANTE ALLA CORNEA NECESSARIO PER SOPPRIMERE LA MELATONINA NOTTURNA DEL 35% PER OGNI PICCO DI LUNGHEZZA D’ONDA (PΛ), TALE VALORE SECONDO BRAINARD E THAPAN SI AGGIRA INTORNO AI 460NM. I DATI RICAVATI DA THAPAN ESPRESSI IN UNITÀ QUANTICHE DI SENSIBILITÀ, SONO STATI ESTRAPOLATI E CONVERTITI IN UNITÀ RADIOMETRICHE. 17
LA SENSIBILITÀ RELATIVA È STATA CALCOLATA MEDIANTE IL RAPPORTO PΛMIN POWER/PΛ; TALI VALORI SONO RIPORTATI IN TABELLA 1 E TRACCIATI IN FIGURA 7 [4]. Brainard et al. (2001, 2008) Thapan et al. (2001) Wavelength (nm) Relative Wavelength Relative sensitivity (nm) sensitivit y 420 (Brainard et al. 0.146 424 0.896 2001) 420 (Brainard et al. 0.219 456 1.000 2008) 440 0.993 474 0.814 460 1.000 494 0.535 480 0.704 520 0.481 Alcuni dei risultati da Brainard e Thapan sono basati sullo stimolo espresso in termini di irradianza (µW/cm2) e altri in quanti (fotoni/cm2/s). La conversione da un’unità ad un’altra è semplice, perciò è possibile combinare le differenti efficienze spettrali stimate, come in Tabella 1 e in Figura 10. Tuttavia, si deve porre molta attenzione poiché la forma della curva dell’efficienza basata sull’irradianza differisce da quella espressa in quanti. Tale artefatto si presenta poiché l’efficienza è definita in termini di lunghezza d’onda richiedente il minor ammontare di potere o quanti per produrre l’effetto della norma la lunghezza d’onda richiedente tale quantità di energia è quella corrispondente al picco della curva dell’efficienza spettrale. Il picco di lunghezza d’onda differirà comunque, a seconda dello stimolo se definito in termini di potere o in quanti. Per tale motivo possono subentrare errori durante la comparazione o la combinazione delle funzioni dell’efficienza spettrale definite con unità radiometriche differenti; la probabilità di tali errori cresce quando le unità radiometriche usate per calcolare l’efficienza spettrale non sono definite in modo esplicito. Brainard pubblicò un’ulteriore funzione dose-risposta a 420 nm e attraverso lo stesso metodo di Rea, è stata misurata la FIGURA 11: ERRORI RESIDUI DELLE QUATTRO PREVISIONI sensibilità spettrale del sistema DELLA SENSIBILITÀ SPETTRALE DEL SISTEMA CIRCADIANO circadiano a tale lunghezza d’onda, ALLE SORGENTI A RISTRETTA BANDA. GLI ERRORI MEDI anch’essa inclusa nella Tabella 1 e in RESIDUI PER IL MODELLO A SINGOLA OSPINA A 460 NM, IL Figura 10. MODELLO MELANOPICO, LA FUNZIONE DI GALL E IL MODELLO CLA SONO RISPETTIVAMENTE: 0.14, 0.23, 0.12 Sia Brainard che Thapan quantificano le E 0.06 [4]. loro rispettive stime usando una singola ipotetica opsina, con un picco di sensibilità a circa 460 nm. La Figura 10 mostra la funzione della sensibilità di una singola opsina corretta per il 18
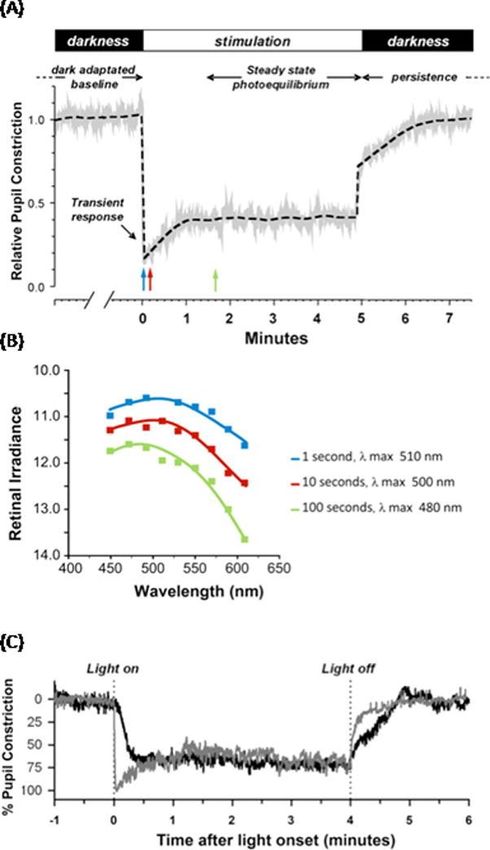
fattore di trasmissione spettrale del cristallino, analoga a quella usata da Brainard e Thapan. La Figura 11 mostra l’errore residuo del modello a singola opsina a 460 nm in funzione della lunghezza d’onda; l’errore residuo è per definizione la differenza assoluta tra la sensibilità misurata ad una data lunghezza d’onda e la sensibilità adattata a quella lunghezza d’onda. Il modello a singola opsina però non appare adeguato per la caratterizzazione del set di dati empirici alle lunghezze d’onda più corte e più lunghe di 460 nm, e inoltre è impreciso dal punto di vista fisiologico poiché non si è a conoscenza di alcun fotopigmento, risiedente nella retina che abbia un picco di sensibilità a tale lunghezza d’onda Con la scoperta della melanopsina, Enezi et al. hanno proposto una funzione della sensibilità melanopica a singola opsina (Figura 10), basata appunto sull’assorbimento spettrale di tale fotopigmento. I dati raccolti sono stati ottenuti attraverso esperimenti su topi geneticamente modificati, ed in base a questi il gruppo di ricercatori sostiene che il riflesso pupillare del topo ad uno stimolo luminoso o ad impulsi di luce intermittenti, può essere spiegato attraverso l'esistenza di una singola opsina che abbia un picco di sensibilità vicino a 480 nm. Tale modello se ponderato per l'assorbimento delle lenti dell'occhio umano, può essere utilizzato per capire la sensibilità spettrale del sistema circadiano dell'uomo. Tale funzione melanopica è inclusa in Figura 1, ma non caratterizzando al meglio i dati ottenuti empiricamente, è associato ad essa il grafico dell'errore residuo. Già prima della scoperta delle ipRGCs e del loro fotopigmento, la melanopsina, vi erano chiari indizi nel grafico della sensibilità spettrale ottenuto empiricamente, del fatto che la fototrasduzione circadiana non può essere predetta dalla sensibilità spettrale di una singola ospina. Innanzitutto, l’inviluppo della sensibilità spettrale definito dalle stime empiriche è più ampio rispetto a quello delle singole opsine; infatti, i modelli a singola opsina in Figura 10 non predicono al meglio la risposta del sistema circadiano dell'uomo allo stesso modo di quella ottenuta mediante la soppressione della melatonina. Inoltre, sia i dati raccolti da Brainard che da Thapan presentano delle discontinuità tra i 470 nm e i 530 nm (Figura 10), suggerendo il coinvolgimento di meccanismi neurali che vedono il contributo di differenti fotopigmenti. In accordo con ciò, Hattar et al. dimostrarono che molteplici fotopigmenti intervengono nella fototrasduzione circadiana nei mammiferi: sebbene le ipRGCs siano i principali fotorecettori, essi ricevono il contributo dai classici coni e bastoncelli. Dato che questi ultimi non possono connettersi direttamente alle ipRGCs, lo fanno attraverso connessioni neurali con cellule bipolari, orizzontali ed amacrine. Per aggirare tale problematica, Gall utilizzò un modello arbitrario per caratterizzare le stime empiriche della sensibilità spettrale di Brainard e Thapan. La funzione di tale modello nonostante sembri esser stata derivata in unità quantiche invece che unità di potere radiante, è simile all’ortodossa funzione dell’efficienza luminosa per il fatto che essa è più vasta rispetto a quella basata su una singola opsina, ma si differenzia perché ignora le discontinuità in entrambi i gruppi di stime tra i 470 nm e i 530 nm (Figura 10). Di conseguenza, utilizzando la funzione di Gall non sarà possibile stimare la sensibilità spettrale a lunghezze d’onda più corte e più lunghe di 460 nm, come mostrato in Figura 11; generalmente, comunque, soddisfa maggiormente l’insieme di stime empiriche della sensibilità spettrale di Brainard e Thapan rispetto a quella a singola opsina [4]. 19
CAPITOLO 2 La scoperta di un nuovo fotorecettore: ipRGC Durante l’ultima decade è stato scoperto che mentre la capacità fotorecettiva della retina è dominata dai coni e bastoncelli, alcuni dei neuroni retinici di uscita (cellule gangliari retiniche) sono anch’essi direttamente fotosensibili. Tali tipi di neuroni, situati prevalentemente nella parte inferiore della retina, sono stati definiti come ipRGC (intrinsecally photosensitive retinal ganglion cells), cellule gangliari intrinsecamente fotosensibili. Le ipRGC comprendono solo una piccola frazione della popolazione totale delle cellule gangliari (1-5%, a seconda della specie e metodo di stima), ma proiettano alle porzioni retino-recettive più estese del cervello, incluse quelle associate alla risposta non-visiva. Difatti, con l’invecchiamento della retina, tale organo subisce modifiche e la visione peggiora a causa dell’ispessimento, dell’ingiallimento e della minore trasmittanza luminosa del cristallino che, provocando una diminuzione di trasparenza dell’occhio alle lunghezze d’onda più corte, influenzano l’ampiezza dei cicli circadiani. Per di più specifiche ablazioni delle ipRGC sopprimono le risposte non-visive, identificando tale classe di cellule come il principale condotto dello stimolo fotogeno alla risposta circadiana e altre risposte sistemiche alla luce. Le ipRGC possono rilevare la luce persino se isolate dal resto della retina, spiegando perché una certa fotosensibilità sopravvive con la perdita delle funzionalità dei coni e bastoncelli, sono da considerare dunque il terzo fotorecettore retinico, il cui compito è quello di catturare “l’informazione non-visiva” della luce e di trasmetterla con lo scopo di attivare il sistema circadiano. Tali recettori però, differiscono in modo significativo per diversi aspetti dai classici coni e bastoncelli. Innanzitutto, si discostano da questi ultimi per l’elemento di fotorecezione, invece di esprimere rodopsina o le opsine dei coni, le ipRGC utilizzano una proteina correlata ma unica, sensibile alla luce, chiamata melanopsina. In secondo luogo, le ipRGC depolarizzano in risposta alla luce, e se lo stimolo è abbastanza forte, generano potenziali d’azione. I coni e i bastoncelli iperpolarizzano quando illuminati e non generano mai potenziali d’azione. In terzo luogo, per raggiungere la soglia di attivazione, le ipRGC richiedono luce molto forte, diversi ordini di grandezza oltre la soglia per le risposte dei coni. In quarto luogo, la risposta delle ipRGC richiede pochi secondi ed esse mantengono fedelmente le loro risposte anche quando i livelli di luce sono mantenuti per ore [22]. 2.1 La risposta delle ipRGC alla luce La melanopsina, il fotopigmento delle ipRGC, è strutturalmente e filogeneticamente più strettamente collegato alle opsine dei fotorecettori rabdomerici degli invertebrati che le opsine dei coni e bastoncelli. In comune con suddette opsine degli invertebrati la cascata di fototrasduzione assunta dalla melanopsina, risulta nella depolarizzazione cellulare. Come risultato, la fondamentale risposta alla luce delle ipRGC dipende dall’irradianza ed accresce nell’attivazione. 20
FIGURA 12: TUTTI LE CLASSI DI FOTORECETTORI RETINALI SONO A MONTE DELLE RISPOSTE CIRCADIANA, NEUROENDOCRINA E NEURO-COMPORTAMENTALE ALLA LUCE. a) SCHEMA DEI CIRCUITI RETINICI DELL’ESSERE UMANO. LE RISPOSTE DI NON FORMAZIONE DELL’IMMAGINE ORIGINANO NELLA RETINA E SONO STATE ATTRIBUITE A UNA PARTICOLARE CLASSE DI CELLULE RETINALI GANGLIONARI (IPRGCS). LE IPRGCS SONO CONNESSE AL CIRCUITO RETINICO CONVENZIONALE TRAMITE I FOTORECETTORI ESTERNI CONI E BASTONCELLI. I DETTAGLI DELLE LORO CONNESSIONI INTRARETINALI NON SONO COMPRESE DEL TUTTO E PROBABILMENTE VARIANO TRA I DIFFERENTI SOTTOTIPI. DALLA FIGURA SI VEDE CHE LE MAGGIORI CONNESSIONI SI HANNO CON LE CELLULE BIPOLARI DEI CONI (CBCS) CONNESSE AI CONI, ALLE CELLULE AMACRINE (ALL) E LE CELLULE BIPOLARI DEI BASTONCELLI (RBC) CONNESSE AI BASTONCELLI. COME CONSEGUENZA, L’ATTIVAZIONE DELLE IPRGCS PUÒ ESSERE INFLUENZATA SIA DALLA FOTORECEZIONE INTRINSECA DELLA MELANOPSINA E I SEGNALI ESTRINSECI CHE ORIGINANO NEI BASTONCELLI E NEI TRE TIPI DI CONI. b) TALE CARATTERISTICA È CONCETTUALIZZATA IN MOLTE FORME SEMPLIFICATE COME UN NUMERO DI MECCANISMI FOTORECETTIVI, OGNUNO DEI QUALI ASSORBE LA LUCE IN ACCORDO AL PROPRIO PROFILO DI SENSIBILITÀ SPETTRALE PER GENERARE UNA DISTINTA MISURA DELL’ILLUMINANZA. TALI CINQUE SEGNALI SONO POI COMBINATI MEDIANTE CONNESSIONI RETINICHE, E CON LE STESSE IPRGC, PER PRODURRE UN SEGNALE INTEGRATO CHE È INVIATO SUCCESSIVAMENTE AI CENTRI DI NON-FORMAZIONE DELL’IMMAGINE DEL CERVELLO. COME OGNUNO DI QUESTE CINQUE RAPPRESENTAZIONI DELL’IRRADIANZA PESATA È PRODOTTA DA UN FOTOPIGMENTO CON UN PROPRIO PROFILO DI SENSIBILITÀ SPETTRALE, LA LORO RELATIVA IMPORTANZA NEL SEGNALE INTEGRATO D’USCITA, DEFINISCE LA DIPENDENZA DALLA LUNGHEZZA D’ONDA DI TALE SEGNALE E LE SUE RISPOSTE A VALLE [22]. L’efficienza quantica della melanopsina è comparabile a quella delle opsine dei coni e bastoncelli. Le ipRGC, comunque, non hanno organelli specializzati dove vi è concentrato il fotopigmento (come quello dei segmenti esterni dei coni e bastoncelli) che massimizzano la probabilità di cattura dei fotoni. Ciò comporta che è la probabilità di assorbire un fotone, per una data area di stimolazione, è 1 milione di volte più bassa rispetto a quella nei coni e bastoncelli. Dunque, nonostante la cascata di fototrasduzione ha un’elevata amplificazione, la fotorecezione della melanopsina è molto meno sensibile rispetto a quella dei coni e bastoncelli. Una volta che è stata raggiunta la soglia di attivazione della melanopsina, comunque, la risposta intrinseca alla luce diminuisce di diversi ordini decimali con l’intensità dello 21
stimolo ed è notevolmente persistente, venendo sostenuto da lunghe durate di illuminazione costante. Sebbene la fototrasduzione della melanopsina è attivata solo da moderate o alte irradianze, le ipRGC e le loro risposte a valle possono essere sensibili a livelli molto più bassi di illuminazione. Per esempio, originariamente si pensava che era richiesto un illuminamento di 2500 lux per la soppressione notturna della melatonina negli esseri umani, ma studi recenti hanno mostrato che sotto certe condizioni, già solo un illuminamento di 1 lux e anche meno può sopprimere la melatonina. Tale sensibilità evidenzia un importante aspetto di tale sistema fotorecettivo: le ipRGC ricevono lo stimolo dalla retina esterna (Figura 12A). Così i dendriti delle ipRGC sono diretti, attraverso stimoli sinaptici alle cellule bipolari ed amacrine, oltre ad essere il sito per la fototrasduzione guidata dalla melanopsina. Come risultato, lo schema di attivazione delle ipRGC è un segnale complesso ed integrato costituito dalla risposta intrinseca alla luce (fotorecezione della melanopsina) e i segnali in entrata provenienti da coni e bastoncelli (Figura 12B). Tale assetto estende considerevolmente intervallo dello stimolo che può suscitare le risposte circadiana e neurofisiologica e spiega perché gli animali che sono geneticamente e privi di melanopsina continuano ad manifestare risposte alla luce di “non formazione dell’immagine” [22]. 2.1.1 Sensibilità spettrale delle ipRGCs Già dalla sua origine, il segnale che guida le risposte fisiologiche e comportamentali alla luce, ovvero l’attivazione delle ipRGCs, è definito dall’influenza combinata di multipli processi fotorecettivi: il meccanismo di fototrasduzione guidato dalla melanopsina all’interno delle stesse ipRGCs, e la fotorecezione remota dei coni e bastoncelli. Ognuno di questi meccanismi di rilevazione della luce ha una distinta sensibilità spettrale, definita dall’efficienza spettrale del fotopigmento espresso e le proprietà di trasmissione spettrale dell’occhio. Le sensibilità spettrali dei coni e bastoncelli sono state già discusse nel precedente capitolo, di seguito verrà analizzata in maggior dettaglio la sensibilità spettrale della melanopsina [22]. 2.1.2 Sensibilità spettrale della melanopsina I dati a disposizione che emergono da ricerche scientifiche indicano che la sensibilità spettrale della melanopsina, il fotopigmento delle ipRGCs, è invariato tra le specie ed ha un picco ad approssimativamente 480 nm. La potenziale complicazione che emerge dalla comparazione di suddette stime della sensibilità spettrale alle proprietà delle risposte spettrali delle ipRGC in vivo, suggerisce che come l’opsina rabdomerica degli invertebrati, la melanopsina potrebbe essere bistabile. La bistabilità permette ai fotopigmenti rabdomerici la capacità di rigenerare il cromoforo derivato dalla vitamina A che è isomerizzato a seguito dell’assorbimento della luce, attraverso l’assorbimento di un altro fotone. Poiché tale rigenerazione può essere prodotto da differenti lunghezze d’onda, potrebbe influenzare le proprietà della risposta spettrale del recettore. 22
Puoi anche leggere