Carboidrati, lipidi, amminoacidi
←
→
Trascrizione del contenuto della pagina
Se il tuo browser non visualizza correttamente la pagina, ti preghiamo di leggere il contenuto della pagina quaggiù
Carboidrati, lipidi,
amminoacidi
1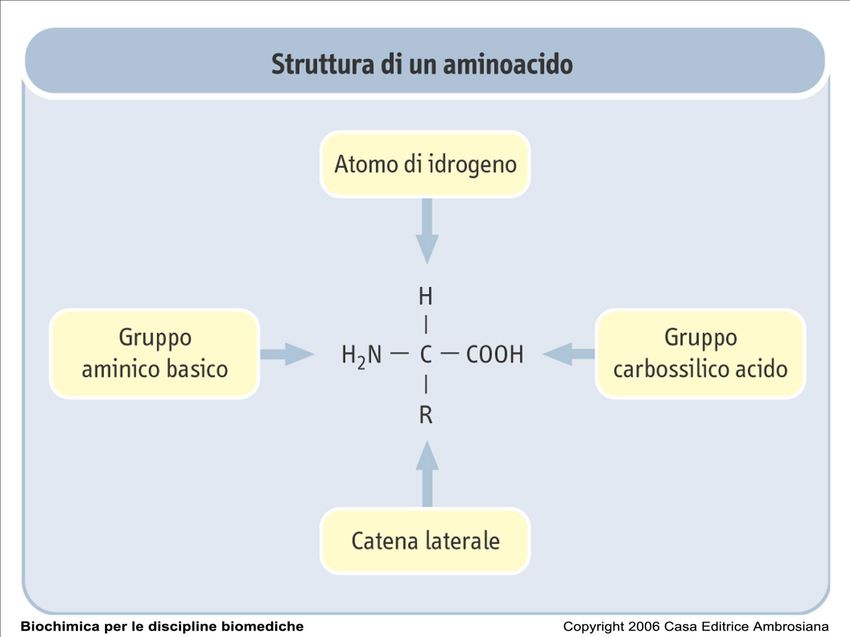
I carboidrati I carboidrati (o saccaridi o glucidi) hanno formula bruta (CH2O)n, dove n è maggiore di 3. Chimicamente sono poli-idrossi chetoni o aldeidi ed esistono sia in forma semplice (monosaccaridi) che come polimeri (oligo- e poli-saccaridi) formati dall'unione di più monosaccaridi. Rappresentano la principale fonte di energia per gli organismi. Il monosaccaride più importante in questo senso (e anche il più abbondante) è il glucosio. Da esso derivano la maggior parte dei polisaccaridi che comprendono importanti sostanze di riserva (amido) o strutturali (cellulosa). © Zanichelli editore, 2014 2
I monosaccaridi (I) I monosaccaridi più semplici hanno 3 atomi di carbonio e sono detti triosi (ad es. gliceraldeide). A seconda del numero di atomi di carbonio si distinguono tetrosi (4), pentosi (5), esosi (6) e così via. I monosaccaridi hanno scheletri non ramificati in cui ad ogni atomo di carbonio, eccetto uno, sono legati gruppi ossidrilici. Il rimanente atomo di carbonio porta legato un ossigeno carbonilico. Se questo è all'estremità della catena (gruppo aldeidico) il saccaride è un aldoso. Se l'ossigeno carbonilico è legato ad un carbonio all'interno della catena (gruppo chetonico) il saccaride è un chetoso. © Zanichelli editore, 2014 3
I monosaccaridi (II)
Gli otto monosaccaridi essenziali per l’uomo
© Zanichelli editore, 2014 4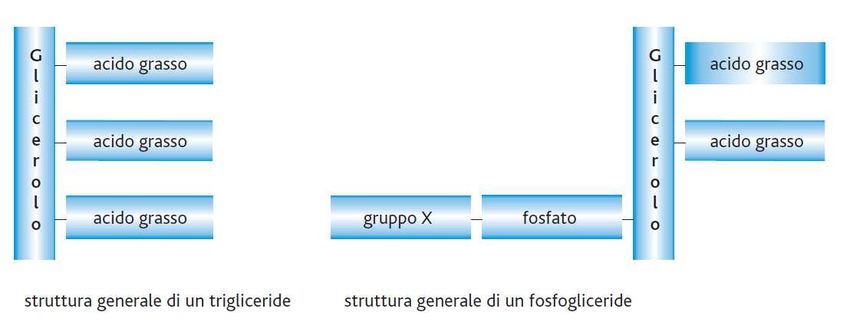
Isomeria ottica D- e L- dei saccaridi (I) I monosaccaridi contengono un centro chirale, ovvero un carbonio legato a quattro sostituenti differenti (detto stereocentro). Questo fa sì che esistano due stereoisomeri. Nel 1891 Emil Fischer introdusse una rappresentazione grafica a croce (proiezione di Fisher) e propose di denominare i due stereoisomeri della gliceraldeide L- gliceraldeide e D-gliceraldeide, sulla base della posizione del gruppo –OH rispetto allo stereocentro. © Zanichelli editore, 2014 5
Isomeria ottica D- e L- dei
saccaridi (II)
Proiezioni di Fischer dei due stereoisomeri della gliceraldeide. L’asterisco indica lo
stereocentro.
© Zanichelli editore, 2014 6Isomeria ottica D- e L- dei saccaridi (III) Prendendo la gliceraldeide come riferimento, è possibile assegnare la configurazione D- o L- a tutti gli altri monosaccaridi, guardando la posizione del gruppo –OH rispetto al carbonio asimmetrico. I monosaccaridi naturali esistono prevalentemente in forma D-. Quattro monosaccaridi di configurazione D-. L’asterisco indica lo stereocentro. © Zanichelli editore, 2014 7
Ciclizzazione (I) Aldeidi e chetoni reagiscono spontaneamente in ambiente acquoso generando emiacetali o emichetali. Quindi, i gruppi alcolici (-OH) dei monosaccaridi tendono a reagire con i gruppi chetonici o aldeidici nella stessa molecola. Questa reazione di ciclizzazione genera una struttura ad anello. In soluzione, i monosaccaridi esistono principalmente (>90%) in questa forma ciclica. Per rappresentare le forme cicliche si utilizza la proiezione di Haworth. © Zanichelli editore, 2014 8
Ciclizzazione (II)
Un esoso ciclico rappresentato con la proiezione di Haworth
© Zanichelli editore, 2014 9Ciclizzazione degli esosi Osserviamo la reazione dei due monosaccaridi più abbondanti: il glucosio (aldoesoso) e il fruttosio (chetososo). © Zanichelli editore, 2014 10
Furanosi e piranosi In entrambi i casi, è il gruppo –OH del carbonio C5 che reagisce con l'ossigeno carbonilico. Nel glucosio questo è in posizione C1 (gruppo aldeidico) per cui la reazione genera un anello a 6 atomi. Nel fruttosio, l'ossigeno carbonilico è in C2 (gruppo chetonico) e la reazione genera un anello a 5 atomi. I monosaccaridi con anello a 6 sono detto furanosi, quelli con anello a 5 sono detti piranosi, per la loro somiglianza con gli eterocicli furano e pirano. © Zanichelli editore, 2014 11
Anomeri Quando i monosaccaridi ciclizzano, il gruppo –OH legato al C1 può trovarsi dallo stesso lato del gruppo CH2OH legato al carbonio 6, rispetto al piano dell'anello (posizione cis), oppure dal lato opposto (posizione trans). Gli isomeri di posizione rispetto al C1 sono detti anomeri. I monosaccaridi con l' –OH al C1 in posizione cis sono detti anomeri β, quelli in trans sono detti anomeri α. © Zanichelli editore, 2014 12
I disaccaridi Il gruppo –OH del carbonio anomerico (C1 nel glucosio e C2 nel fruttosio) di un monosaccaride ciclico, reagisce con gli alcoli a formare un acetale. Se il gruppo alcolico –OH che reagisce con il gruppo idrossile del carbonio anomerico è quello di un altro monosaccaride, il prodotto è un disaccaride: cioè due monosaccaridi legati tra di loro da un legame glicosidico in cui un ossigeno fa da ponte tra i due anelli. I disaccaridi sono quindi acetali, in cui l' –OH anomerico diventa un gruppo –OR, dove R rappresenta il secondo monosaccaride. L'ossigeno del legame glicosidico può presentarsi nelle due forme anomeriche α o β. © Zanichelli editore, 2014 13
Il maltosio Il maltosio (α-D-glucopiranosil-(1->4)-α-D glucopiranoso) è formato da due residui di α-D-glucosio legati con legame glicosidico 1 -> 4 (cioè il gruppo –OH anomerico in C1 reagice con quello alcolico in C4 della seconda molecola di glucosio). Il legame glicosidico rispetto al carbonio anomerico è di tipo α. Il maltosio è un prodotto della degradazione dell'amido. © Zanichelli editore, 2014 14
Il cellobioso Lo stesso monosaccaride può originare disaccaridi differenti in base alla sua configurazione anomerica. Ad esempio, se il D-glucosio è di tipo β, l'unione di due residui di β-D-gIucosio dà il cellobioso (β-D-glucopiranosil-(1-> 4)-β-D glucopiranoso). Il legame glicosidico del cellobioso è 1 -> 4 come nel maltosio, ma la configurazione rispetto al carbonio anomerico è di tipo β. Il cellobioso è un prodotto della degradazione della cellulosa. © Zanichelli editore, 2014 15
Il lattosio Il lattosio(β-D-galattopiranosil-(1->4)-β-D- glucopiranoso) è formato da un residuo di β-D-galattosio e uno di α-D-glucosio legati con legame glicosidico 1 -> 4. Il legame glicosidico rispetto al carbonio anomerico è di tipo β. Il lattosio si trova unicamente nel latte. © Zanichelli editore, 2014 16
Il saccarosio Il saccarosio (β-D-fruttofuranosil-(2->1)-α-D- glucopiranoside) è formato da un residuo di β-D- fruttosio e uno di α-D-glucosio. È un disaccaride particolare in quanto il legame glicosidico coinvolge entrambi i carboni anomerici (C2 del fruttosio e C1 del glucosio). Il saccarosio è lo zucchero di canna e di barbabietola. © Zanichelli editore, 2014 17
I polisaccaridi In natura, i carboidrati si trovano sotto forma di polisaccaridi, ovvero lunghi polimeri formati dall'unione di centinaia o migliaia di unità monosaccaridiche attraverso il legame glicosidico. Si dicono omopolisaccaridi quelli formati da un unico tipo di monosaccaride ed eteropolisaccaridi quelli formati da monosaccaridi di tipo diverso. I polisaccaridi più importanti biologicamente sono classificabili come polisaccaridi di deposito (es. amido, glicogeno) e strutturali (es. cellulosa). © Zanichelli editore, 2014 18
Amido L'amido è un omopolisaccaride formato da residui di α-D-glucosio. Esiste in due forme: una lineare, α- amilosio, in cui il legame glicosidico è di tipo α 1-> 4 e una ramificata, amilopectina, in cui catene lineari α 1-> 4 sono collegate tra di loro con legami di tipo α 1-> 6. L'amido è il principale polisaccaride di deposito nelle piante. © Zanichelli editore, 2014 19
Glicogeno Il glicogeno è un omopolisaccaride formato da residui di α-D-glucosio. Ha una struttura altamente ramificata in cui le catene lineari α 1-> 4 si ramificano con legame di tipo α 1-> 6 ogni 8-12 residui. Il glicogeno è il principale polisaccaride di deposito nelle cellule animali. Rappresenta fino al 10% del peso del fegato e l'1-2% del peso del muscolo scheletrico. © Zanichelli editore, 2014 20
Cellulosa La cellulosa è un omopolisaccaride formato da residui di β-D-glucosio. La sua struttura è lineare, del tutto analoga a quella dell' α-amilosio, ma con l'importante differenza che la configurazione anomerica del legame glicosidico è di tipo β. La cellulosa è il polisaccaride più abbondante in natura e costituisce il principale componente strutturale del mondo vegetale. © Zanichelli editore, 2014 21
I LIPIDI
I lipidi I lipidi sono complesse molecole organiche insolubili in acqua che svolgono importanti funzioni: • riserva energetica; • ruolo strutturale delle membrane cellulari; • ruoli funzionali (ormoni, vitamine). Si possono distinguere in lipidi complessi che contengono acidi grassi e lipidi semplici che ne sono privi. I lipidi complessi sono detti anche saponificabili, in quanto gli acidi grassi presenti possono andare incontro a idrolisi in ambiente basico, reazione detta di saponificazione e che genera i saponi (sali di acidi grassi). © Zanichelli editore, 2014 23
Gli acidi grassi Gli acidi grassi sono costituiti da lunghe catene idrocarburiche terminanti con un gruppo carbossilico: CH3(-CH2-)n-COOH. Possono essere saturi (cioè gli atomi di carbonio sono collegati da legami singoli) o insaturi (cioè possono essere presenti uno o più doppi legami). Gli acidi grassi saturi più comuni sono l'acido palmitico (16 atomi di carbonio) e l'acido stearico (18 atomi di carbonio). L'acido insaturo più comune è l'acido oleico (18 atomi di carbonio e un doppio legame tra C9 e C10). © Zanichelli editore, 2014 24
I lipidi complessi (I) I lipidi complessi contengono acidi grassi e comprendono i trigliceridi, i fosfogliceridi, gli sfingolipidi e le cere. • I trigliceridi o triacilgliceroli sono esteri degli acidi grassi e del glicerolo, in cui tutti e tre i gruppi –OH del glicerolo sono esterificati. • I fosfogliceridi o fosfolipidi sono esteri del glicerolo, in due –OH sono esterificati da acidi grassi, mentre al terzo –OH è legato un gruppo fosforico a sua volta legato ad un gruppo X che può essere di varia natura (ad es. l'amminoacido serina o un alcol come il glicerolo). Questa estremità è detta testa polare. © Zanichelli editore, 2014 25
I lipidi complessi (II)
• Gli sfingolipidi sono composti dalla sfingosina
legata con legame amidico ad un acido grasso.
• Le cere sono esteri solidi di acidi grassi complessi
con alcoli a lunga catena.
Alcuni tipi comuni di lipidi di riserva e di membrana.
© Zanichelli editore, 2014 26Trigliceridi e fosfolipidi I trigliceridi sono i principali componenti dei lipidi di deposito delle cellule animali. I fosfolipidi (o fosfogliceridi) sono i principali componenti delle membrane cellulari, in cui due strati di fosfolipidi si affacciano l'uno verso l'altro con le code idrofobiche, esponendo le teste polari verso l'ambiente acquoso extracellulare e quello citoplasmatico. © Zanichelli editore, 2014 27
Lipidi semplici: i terpeni I lipidi semplici non contengono acidi grassi e sono quindi insaponificabili. Esistono due classi principali di lipidi semplici, i terpeni e gli steroidi. • I terpeni sono costituiti da multiple unità di isoprene (2 metil, 1,3 butadiene) e possono essere ciclici e lineari. Tra i terpeni più comuni vi sono oli essenziali profumati come la canfora o il mentolo, le vitamine A, E, K e il β-carotene. Struttura dell’isoprene, unità di base dei terpeni © Zanichelli editore, 2014 28
Lipidi semplici: gli steroidi
Gli steroidi hanno come nucleo un idrocarburo
tetraciclico, il ciclopentanoperidroferantrene,
formato da tre anelli a 6 atomi di carbonio e uno a 5,
fusi insieme. La sintesi degli steroidi nella cellula parte
dal composto terpenico squalene, da cui derivano il
lanosterolo e il colesterolo. Dal colesterolo derivano
poi altre biomolecole essenziali come gli acidi biliari e
gli ormoni androgeni e estrogeni e il progesterone.
Struttura del
ciclopentanoperidroferantrene
© Zanichelli editore, 2014 29Lipidi semplici: esempi
Colesterolo
Vitamina A (tutto-trans retinolo)
30GLI AMMINOACIDI
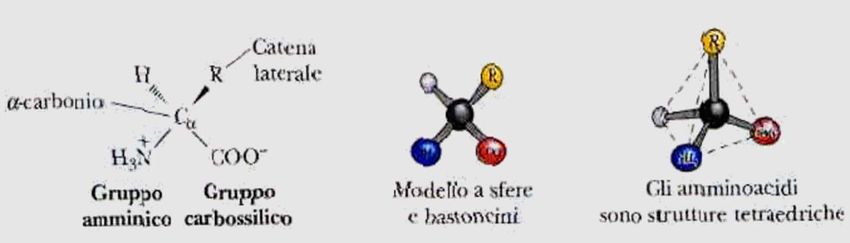
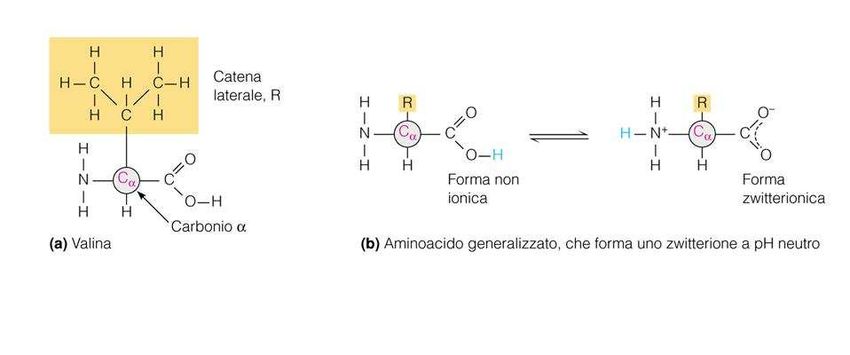
Gli amminoacidi (I)
Gli amminoacidi sono molecole formate da un gruppo
amminico (-NH2) e un gruppo carbossilico (-COOH)
legati ad un carbonio centrale detto carbonio α.
Al Cα sono legati un idrogeno e un gruppo R, detto
anche catena laterale o gruppo funzionale,
caratteristico di ogni amminoacido e che ne determina
le proprietà chimico-fisiche.
Nell'amminoacido glicina, R è un altro atomo di
idrogeno, per cui il Cα non è un centro asimmetrico. In
tutti gli altri amminoacidi, Cα è legato a quattro gruppi
diversi, per cui determina un centro chirale.
32Gli amminoacidi (II)
Sebbene possano esistere entrambi gli isomeri D- e L-,
in natura tutti gli amminoacidi sono L- (ad eccezione
di alcuni amminoacidi liberi non proteici, presenti nella
parete batterica).
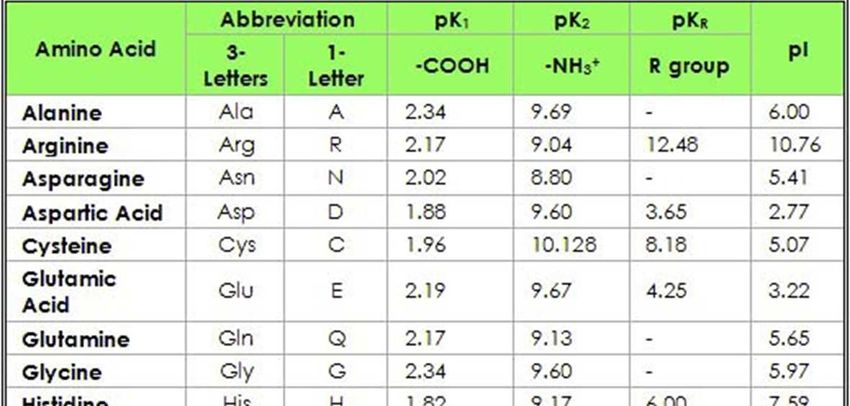
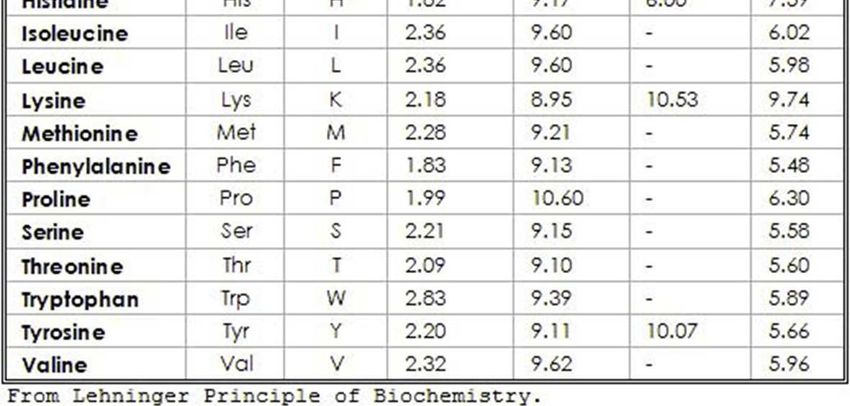
34Proprietà acido-basiche degli amminoacidi
(I): molto importanti per comprendere e analizzare le proteine
Poiché nella molecola degli AA sono presenti gruppi –NH2 e –COOH, basici
deboli e acidi deboli, questi possono ionizzarsi in risposta a variazioni di
pH del mezzo acquoso.
Questo spiega perché gli AA hanno punti di fusione abbastanza alti e sono
molto solubili nei solventi polari
35ZWITTERIONE
• Quando un amminoacido viene sciolto in H2O diventa
uno ione dipolare (zwitterione) che può agire sia come
acido (donatore di protoni) che come base (accettore di
protoni)
• Le sostanze che hanno questa doppia natura si
definiscono anfòtere o anfoliti.
• Al pH fisiologico (valore attorno a 7,4) tutti gli
amminoacidi hanno:
– il gruppo carbossilico dissociato,si forma lo ione negativo
carbossilato -COO-)
– il gruppo amminico protonato (-NH3+)Proprietà acido-basiche degli
amminoacidi (II)
Il gruppo -NH2 può accettare un protone, mentre il
gruppo –COOH può cederlo: questa caratteristica
rende gli amminoacidi degli anfoliti o zwitterioni, cioè
ioni dipolari in grado di comportarsi sia come acidi
che come basi.
37catione zwitterione anione
CURVA DI TITOLAZIONE ALANINA A pH =6,02 si ha il pI, Punto isoelettrico al quale non vi è alcuna carica netta. Ciascun AA ha un proprio punto isoelettrico.
pH>pKa = Protone dissociato
pHProprietà chimico-fisiche degli amminoacidi (I) Oltre alla parte funzionale, comune a tutti, ogni amminoacido presenta un gruppo -R proprio La natura del gruppo -R conferisce proprietà diverse a ciascun amminoacido. Gli amminoacidi possono essere classificati in base alle proprietà delle loro catene laterali (-R), considerando la loro polarità o non polarità al pH fisiologico e quindi la tendenza ad interagire con l’acqua Gli amminoacidi con catene laterali cariche, idrofiliche, sono generalmente esposti sulla superficie delle proteine I residui idrofobici, non polari, si trovano in genere all’interno delle proteine, protetti dal contatto con l’acqua 42
• Esistono 20 amminoacidi comuni e alcuni amminoacidi rari che costituiscono le unità base delle proteine. • Inoltre esistono oltre 150 amminoacidi non proteici, presenti soprattutto nelle piante e nei funghi.
Proprietà chimico-fisiche degli
amminoacidi (II)
Gli amminoacidi
comuni a tutte le
proteine sono
classificati in base
alle caratteristiche dei
loro gruppi R: 9
apolari (idrofobici); 6
neutri; 3 carica
positiva (basici); 2
carica negativa
(acidi).
45Proprietà chimico-fisiche degli
amminoacidi (III)
Gli amminoacidi
comuni a tutte le
proteine sono
classificati in base
alle caratteristiche dei
loro gruppi R: 9
apolari (idrofobici); 6
neutri; 3 carica
positiva (basici); 2
carica negativa
(acidi).
46Amminoacidi essenziali
L'organismo è in grado di sintetizzare da sé molti
amminoacidi, ma non tutti. Esistono infatti 8
amminoacidi detti essenziali, perché devono
essere introdotti con la dieta. Essi sono:
• fenilalanina
• isoleucina
• lisina
• leucina
• metionina
• treonina
• triptofano
• valina
© Zanichelli editore, 2014 47Puoi anche leggere

















































