UNIVERSITÀ DEGLI STUDI DI NAPOLI "FEDERICO II"
←
→
Trascrizione del contenuto della pagina
Se il tuo browser non visualizza correttamente la pagina, ti preghiamo di leggere il contenuto della pagina quaggiù
UNIVERSITÀ DEGLI STUDI DI NAPOLI
“FEDERICO II”
Scuola Politecnica e delle Scienze di Base
Area Didattica di Scienze Matematiche Fisiche e Naturali
Dipartimento di Fisica “Ettore Pancini”
Laurea Triennale in Fisica
Il pettirosso di Schrödinger:
Interpretazione quantomeccanica della magnetoricezione
Relatori: Candidato:
Prof. Gennaro Miele Antonio Junior Iovino
Matr. N 85000979
Anno Accademico 2018/2019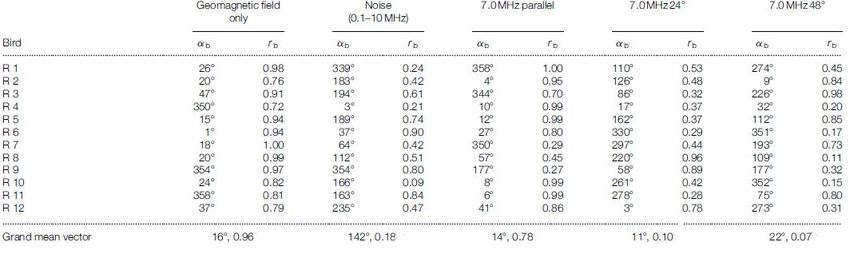
A mia nonna...
"Migrano gli uccelli emigrano
con il cambio di stagione
giochi di aperture alari
che nascondono segreti
di questo sistema solare"
F. Battiato "Gli uccelli"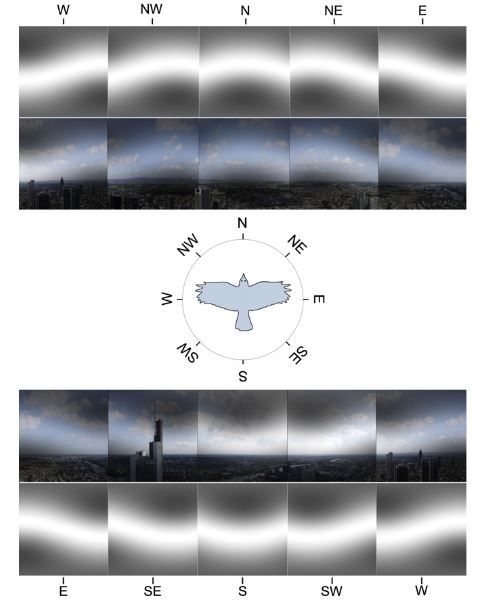
Indice
Introduzione 1
1 La magnetoricezione 2
1.1 La bussola aviaria . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2
1.1.1 Caratteristiche della magnetoricezione . . . . . . . . . . . . . . . . 3
1.2 Meccanismo di coppia di radicali . . . . . . . . . . . . . . . . . . . . . . . 3
1.3 Modello teorico per la magnetoricezione . . . . . . . . . . . . . . . . . . . 5
1.3.1 Triplet yield . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 5
1.3.2 Costruzione dell’Hamiltoniana . . . . . . . . . . . . . . . . . . . . 8
1.3.3 Risultati della simulazione . . . . . . . . . . . . . . . . . . . . . . . 9
2 Visione Magnetica 13
2.1 Modello di bussola aviaria basato sulla visione . . . . . . . . . . . . . . . . 13
2.1.1 Prototipo dell’occhio . . . . . . . . . . . . . . . . . . . . . . . . . . 13
2.1.2 Schemi di risposta . . . . . . . . . . . . . . . . . . . . . . . . . . . 14
2.2 I candidati magnetorecettori . . . . . . . . . . . . . . . . . . . . . . . . . . 18
2.2.1 La magnetite . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 18
2.2.2 I crittocromi . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 19
3 Verifiche sperimentali 21
3.1 L’esperimento di Ritz . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 21
3.1.1 Modello matematico . . . . . . . . . . . . . . . . . . . . . . . . . . 21
3.1.2 Apparato strumentale e procedura del test . . . . . . . . . . . . . . 22
3.1.3 Analisi dei dati . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 23
3.1.4 Risultati . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 25
3.2 Aspetti irrisolti . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 25
3.2.1 La decoerenza nel processo aviario . . . . . . . . . . . . . . . . . . 26
Conclusioni 28
A Il test di Rayleigh 29
Riferimenti bibliografici 29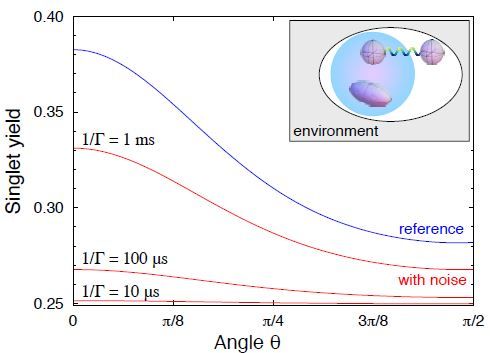
Introduzione
Per secoli il sistema usato dagli animali per orientarsi sul globo è rimasto misterioso. Ora
sappiamo che utilizzano una moltitudine di metodi diversi: alcuni osservano la posizio-
ne del Sole durante il giorno, altri memorizzano punti di riferimento nel paesaggio, altri
ancora, guidati dall’olfatto, annusano letteralmente la via da seguire. Ma è il pettirosso
(Erithacus rubecula) ad avere il sistema di navigazione più misterioso: tale animale ha
la capacità, chiamata magnetoricezione, di rilevare la direzione e l’intensità del campo
magnetico terrestre, e di inferire informazioni sulla direzione da seguire per mezzo di un
senso di navigazione innato, che nel suo caso comporta un nuovo tipo di bussola chimica.
La magnetoricezione rappresenta ancora oggi un enigma nel mondo scientifico; in effetti il
campo magnetico terrestre, il quale varia tra 0,2 e 0,7 Gauss in superficie, è troppo debole
per essere rilevato, nel significato chimico del termine, da un animale, ovvero l’interazione
del campo con le molecole all’interno delle cellule viventi fornisce una quantità di energia
che è meno di un miliardesimo di quella necessaria per creare o distruggere un legame
chimico. E allora, come fa il pettirosso a «percepire» il campo magnetico terrestre?
L’impossibilità di trovare meccanismi classici che potessero spiegare tale capacità con-
dusse la comunità scientifica a ricercare una risposta a tale dilemma all’interno di una
delle teorie scientifiche più sorprendenti del nostro tempo, la meccanica quantistica.
Dopo aver introdotto le caratteristiche principali della Magnetoricezione, la quale si pre-
senta a tutti gli effetti come una sorta di sesto senso, illustrando come tale capacità possa
essere rappresentata da una sorta di bussola magnetica, detta «bussola aviaria», e il per-
ché si possa basare sul meccanismo di coppia di radicali, studieremo un modello teorico
per un organo di senso magnetico basato sulla visione che connette le proprietà molecolari
ai processi quantici della coppia di radicali come risposta comportamentale dell’animale,
ove si propone i criptocromi, una classe di proteine fotosensibili, come candidati per la
bussola chimica; e si descrive in che modo, esattamente, l’orientazione dell’uccello rispetto
al campo magnetico terrestre possa influenzare ciò che vede.
In seguito presenteremo un esperimento, ideato dall’équipe di Ritz all’università di Fran-
coforte, il quale verifica la previsione teorica della coppia di radicali, studiando come
reagisce la bussola del pettirosso in presenza di campi magnetici oscillanti a frequenze
distinte.
Infine discuteremo, in modo discorsivo, in quanto richiamare tutti gli strumenti matema-
tici utili all’analisi del problema ci allontanerebbero troppo dal tema centrale della tesi,
di come il processo alla base della magnetoricezione possa sopravvivere alla decoerenza.
1
Capitolo 1
La magnetoricezione
1.1 La bussola aviaria
Negli anni ’40, studiando la capacità degli uccelli migratori di trovare sempre la via di
casa con tale affidabilità, per ricavare informazioni per uso bellico, H.L. Yeagley e F.C.
Whitmore svilupparono una teoria[20] secondo cui questi animali possono sentire il campo
magnetico terrestre; questo crea una "mappa" nel cervello dell’uccello, che gli fornisce la
latitudine e la longitudine.
Per ottenere delle prove sperimentali concrete a sostegno di questa teoria però bisogna
aspettare il 1965 quando W. Wiltschko[25] iniziò a studiare la capacità di orientamento
dei pettirossi posti in un campo magnetico variabile. Mise i pettirossi, catturati nel mezzo
della migrazione, all’interno di camere magneticamente schermate e li espose a un campo
magnetico artificiale prodotto mediante bobine di Helmholtz, in modo da simulare il
campo geomagnetico e allo stesso tempo cambiare direzione o intensità. Si osservò che
gli uccelli catturati durante la migrazione in autunno o in primavera si agitavano e si
ammassavano nell’angolo della camera che corrispondeva alla direzione di migrazione del
campo magnetico artificiale.
Sempre Wiltschko[41], insieme alla moglie, alcuni anni dopo, nel 1972 ripetendo lo stesso
esperimento, osservò che invertendo la polarità del campo, ruotando un magnete di un
angolo piatto, non si osservava alcun effetto sul comportamento degli uccelli, i quali
continuavano ad orientarsi verso il polo più vicino, qualunque fosse, dimostrando che il
loro senso magnetico non era collegabile ad una bussola classica, bensì ad una bussola ad
inclinazione; questa bussola venne definita bussola aviaria.
Una bussola ad inclinazione è fatta nel seguente modo: visualizziamo la terra come un
enorme magnete con le linee di campo che escono dal polo sud e si irradiano verso l’esterno,
curvandosi per rientrare nel polo nord. La direzione di queste linee in prossimità di ogni
polo è quasi verticale rispetto al suolo, ma esse diventano parallele alla superficie del
pianeta avvicinandosi all’Equatore; una bussola ad inclinazione è in grado di misurare
l’angolo tra le linee di campo magnetico e la superficie terrestre, riuscendo a distinguere
un polo dall’equatore, ma non i due poli come nel caso di una bussola classica.
21 – La magnetoricezione
1.1.1 Caratteristiche della magnetoricezione
Oltre l’essere visualizzabile come una bussola ad inclinazione, negli anni grazie alla sco-
perta della presenza di questo senso magnetico in diverse specie di uccelli[47], si è giunti
all’osservazione di diverse caratteristiche della magnetoricezione, in seguito riportate.
Dipendenza dalla lunghezza d’onda della luce
L’orientazione della bussola dipende dalla lunghezza d’onda della luce ambientale: si è
osservato[51][48] che i pettirossi mostrano un ottimo orientamento in presenza di luce
blu o verde, ma appaiono fortemente disorientati in presenza di luce rossa o gialla, ed è
per questo motivo che le migrazioni di tale animale avvengono principalmente nelle ore
notturne.
Questi studi suggeriscono, inoltre, che affinché il meccanismo della bussola funzioni, la
luce deve presentare un’energia superiore ad una certa soglia.
Dipendenza dall’intensità del campo magnetico
Un’ulteriore importante proprietà è la sensibilità limitata della bussola ad un piccolo
range di intensità di campo magnetico: si è osservato[50] che i pettirossi riescono ad
orientarsi bene soltanto in presenza di cambi magnetici con intensità comprese tra i 0,3
e i 0,7 Gauss, mentre risultano disorientati in presenza di campi con intensità diverse.
1.2 Meccanismo di coppia di radicali
Il chimico tedesco Klaus Schulten, propose verso la fine degli anni ’70 un meccanismo,
chiamato meccanismo di coppia di radicali, per cui sia i campi magnetici che la luce
riescono a influenzare le reazioni chimiche, e quindi nel nostro caso, fornire una bussola
magnetica agli uccelli.
Oggi, tale meccanismo rappresenta l’ipotesi di maggiore spessore su cui si basa il funzio-
namento della bussola aviaria.[36] [35]
In chimica, si definisce radicale[37] (o radicale libero) un’entità molecolare molto reattiva
avente vita media di norma brevissima, costituita da un atomo o una molecola formata da
più atomi, che presenta un elettrone spaiato: tale elettrone rende il radicale estremamente
reattivo, in grado di legarsi ad altri radicali o di sottrarre un elettrone ad altre molecole
vicine.
Il meccanismo di coppia di radicali è un processo intermedio di una reazione in cui si
forma simultaneamente una coppia di radicali; gli elettroni spaiati della coppia sono in
correlazione, dunque il sistema potrà trovarsi soltanto in uno stato di singoletto o di tri-
pletto.
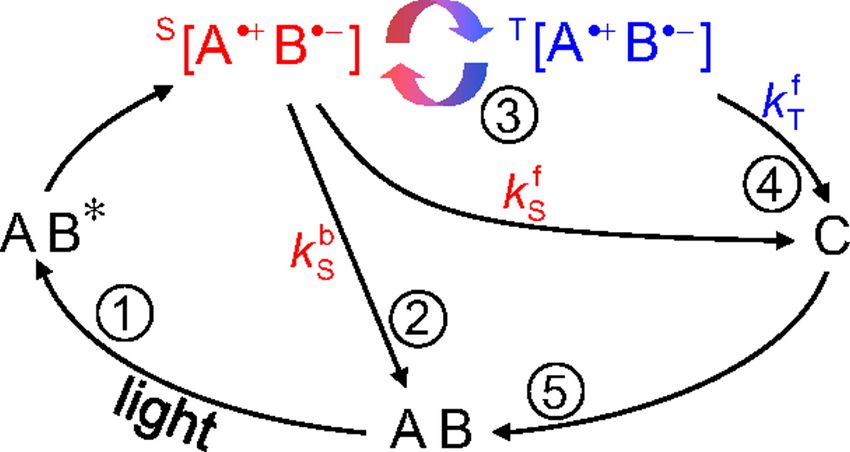
Il meccanismo della coppia di radicali si può così esemplificare (Si veda figura 1.1):
1. Si consideri una generico composto organico AB. Supponiamo si trovi in uno stato
elettronico di singoletto, dato che la maggior parte delle specie organiche allo stato
fondamentale è in questo stato, e che venga eccitato mediante fotoeccitazione; si
31 – La magnetoricezione
crea uno stato intermedio eccitato indicato con AB* che produce una coppia di
radicali indicata con S [A•+ B•- ] , e siccome si conserva lo spin, anche questa coppia
sarà in uno stato di singoletto.
2. Il singoletto S [A•+ B•- ] può reagire in due modi con conservazione dello spin: può
formare un prodotto generico in uno stato di singoletto indicato con C con una
costante di velocità kf s , o perdere l’energia in eccesso e tornare alla specie iniziale
AB con una costante di decadimento kb s .
3. Nel frattempo, siccome lo spin degli elettroni spaiati nella coppia di radicale è ac-
coppiato allo spin nucleare presente dei radicali (interazione iperfine), oltre alla
presenza di un’eventuale campo magnetico esterno (interazione di Zeeman), lo sta-
to di singoletto risulta essere uno stato non-stazionario.
Di conseguenza, si stabilisce un equilibrio tra il singoletto S [A•+ B•- ] e il corrispon-
dente stato di tripletto T [A•+ B•- ].
4. Infine a causa della conservazione dello spin, lo stato di tripletto T [A•+ B•- ] non
può reagire direttamente per formare nuovamente il prodotto finale di reazione C
in stato di singoletto, ciò comporta che la costante di decadimento kf T sarà molto
più piccola di kb s .
Figura 1.1: Schema di meccanismo di coppia di radicali. Fonte:[33]
Allora è lecito pensare che la coppia di radicali in stato di tripletto formi prima dei prodot-
ti intermedi in stato di tripletto(Figura 1.2), con costante di decadimento kT , che vivano
a lungo quasi quanto i prodotti di reazione ottenuti a partire dalla coppia di radicali in
stato di singoletto; ed è proprio questa diversità dei prodotti di reazione ottenuti a causa
della diversa distribuzione di coppie di radicali in stato di singoletto e tripletto a causare
il fenomeno della magnetoricezione.
Ovviamente la probabilità che il meccanismo segua il processo di ricombinazione di stato
di singoletto (tripletto) è proporzionale alla probabilità che la coppia di radicali inizial-
mente si trovi nello stato di singoletto (tripletto).
Il punto chiave del meccanismo della coppia di radicali è che trovandosi in uno stato di
non-equilibrio, la sua dinamica, ovvero l’interconversione singoletto↔tripletto, può esse-
re modificata dalla presenza di un campo magnetico esterno, eventualmente anche molto
debole.
41 – La magnetoricezione
Di conseguenza anche le interazioni magnetiche, pur essendo ordini di grandezza più pic-
cole delle energie di attivazione tipiche dei processi termici, possono influenzare le rese
dei prodotti finali.
Risulta essere particolarmente interessante osservare che se le interazioni interne (in tal
caso l’accoppiamento iperfine) sono anisotropiche, l’effetto di un campo magnetico ester-
no sarà a sua volta anisotropico; ovvero, così come il funzionamento della bussola aviaria
si basa sulla direzione del campo geomagnetico, ugualmente sarà per l’interconversione
tra stato di singoletto e tripletto.
In tale meccanismo, inoltre, si suppone che l’accoppiamento dipolare tra gli elettroni sia
molto più debole rispetto all’accoppiamento iperfine, e che le reazioni di ricombinazione
sono lente abbastanza da poter permettere l’interconversione singoletto↔tripletto.
Figura 1.2: Schema di meccanismo di coppia di radicali, in cui si mostra come si ottengono
prodotti di reazione diversi T P e S P con costanti di decadimento kT e kS . Fonte:[15]
1.3 Modello teorico per la magnetoricezione
1.3.1 Triplet yield
Per valutare quantitativamente l’effetto di un campo magnetico esterno sul meccanismo
di coppia di radicali, introduciamo la grandezza chiamata Triplet yield[8].
La triplet yield ΦT è definita come il rapporto tra la quantità di prodotti di reazione
ottenuta mediante il decadimento di tripletti, e il numero di prodotti di reazione totali.
Per valutare la triplet yield della coppia di radicali, assumiamo che la coppia sia geome-
tricamente fissata in un substrato e che si trovi inizialmente in uno stato di singoletto. In
formule: Z ∞ Z ∞
ΦT = kT T (t) dt = 1 − ΦS = 1 − kS S(t) dt (1.1)
0 0
Dove T(t) e S(t) sono, rispettivamente, la frazione di coppie di radicali in stato di tripletto
e singoletto ad un certo tempo fissato.
Definiti nel seguente modo gli operatori di proiezione di singoletto e tripletto :
1
Q̂S = − Ŝ1 · Ŝ2
4 (1.2)
3
Q̂T = + Ŝ1 · Ŝ2
4
51 – La magnetoricezione
T(t) e S(t) possono essere calcolate nel seguente modo: [12]
S(t) = Tr [QS ρ(t)] (1.3)
T (t) = Tr [QT ρ(t)] (1.4)
dove TrA = i Aii , mentre ρ(t) è la matrice densità.
P
Per poter calcolare la 1.4, possiamo studiare l’evoluzione temporale della matrice di den-
sità ρ(t).
Questo approccio è generalmente usato quando è necessario descrivere un ensamble sta-
tistico di stati, piuttosto che un sistema formato da un solo stato ben definito.[10]
Sebbene possiamo supporre che inizialmente lo stato elettronico di spin sia di singoletto,
lo spin relativo ai nuclei non è noto. Perciò la nostra matrice di densità è descritta da uno
stato elettronico puro di singoletto e da uno stato completamente misto di spin nucleare,
il quale è un ensamble di tutte le possibili configurazioni di spin nucleare.
In formule la matrice di densità può così essere rappresentata:[24]
1
ρ̂(0) = Q̂S ⊗ IZ (1.5)
e Z
Dove Q̂S è l’operatore di proiezione di singoletto nel sottospazio elettronico, IZ è l’opera-
tore identità nel sottospazio relativo allo spin nucleare e 1/Z è il fattore di normalizzazione
riguardante la molteplicità degli stati di spin nucleare:
Z= (2Ii + 1) (1.6)
Y
i
Dove Ii è il numero quantico angolare di spin del i-esimo nucleo. La dimensione dello
spazio formato dai sottospazi nucleari elettronici di spin risulterà dunque essere pari a
4Z.
Per conoscere la forma della matrice di densità ad un generico tempo t, dato lo stato
iniziale, ricorriamo all’equazione di Liouville-von Neumann:
d
ρ̂(t) = −i[Ĥ, ρ̂(t)] − {K̂, ρ̂(t)} (1.7)
dt
Dove [Â, B̂] è il commutatore di  e B̂ mentre {Â, B̂} è l’anticommutatore di  e B̂.
Con Ĥ si rappresenta l’Hamiltoniana di spin della coppia di radicali, mentre K̂ è l’ope-
ratore di ricombinazione di Haberkorn[11] ed è così definito:
kS kT
K̂ = Q̂S + Q̂T (1.8)
2 2
Allora l’equazione 1.7 diventa:
i kS n ˆ o k n
QˆT , ρ(t)
o
T
ρ̇(t) = − [H, ρ(t)] − QS , ρ(t) − (1.9)
~ 2 2
61 – La magnetoricezione
I tre termini che compaiono nell’equazione 1.9 rappresentano, rispettivamente, l’intercon-
versione singoletto↔tripletto nella coppia di radicali causata dall’Hamiltoniana di spin
H, il decadimento della coppia in stato di singoletto nei prodotti di reazione, e il decadi-
mento della coppia in stato di tripletto nei prodotti di reazione.
Un importante semplificazione[42] può essere fatta ponendo ks = kT = k; questa sempli-
ficazione è giustificata dal fatto che se si presentasse una grande differenza tra le costanti
di decadimento, si rimuoverebbe qualsiasi tipo di anisotropia nella resa dei prodotti e
ciò non corrisponderebbe alle caratteristiche possedute dalla bussola aviaria. In effetti se
un modo di decadere risultasse essere dominante sull’altro, quasi la totalità dei prodotti
risulterebbe provenire da quel modo di decadimento, senza tener appunto conto della
direzione del campo magnetico debole esterno.
Questa semplificazione ci permette di scrivere la 1.8 nel seguente modo:
k k k
K̂ = Q̂S + Q̂T = I (1.10)
2 2 2
Ovvero K̂ risulta essere proporzionale all’operatore identità in quanto la somma degli
operatori di proiezione di singoletto e tripletto è la somma su un set completo di operatori
di proiezione.[23]
In presenza di una Hamiltoniana indipendente dal tempo, la matrice di densità, nota la
sua forma all’istante di tempo iniziale, espressa nell’equazione 1.5, può essere espressa nel
seguente modo:[34]
ρ̂(t) = U (t; 0)ρ̂(0)U (t; 0)† = e(−iĤt−K̂t) ρ̂(0)e(+iĤt−K̂t) (1.11)
Dove U (t; 0) rappresenta l’operatore evoluzione temporale.
Tenuto conto che l’operatore identità commuta con ogni operatore, e perciò i termini di
ricombinazione possono essere separati dai termini di interazione nell’equazione 1.11, si
ottiene:
1
ρ̂(t) = e(−iĤt) Q̂S e(+iĤt) e−kIt (1.12)
Z
Siccome l’operatore identità è diagonale, possiamo scrivere l’esponenziale nel seguente
modo e−kIt = Ie−kt .
Andando a calcolare la traccia dell’equazione 1.4 si ottiene:
4N X4N
1 −kt X
T (t) = e · mn Qmn cos [(wm − wn ) t]
QT S
(1.13)
Z m=1 n=1
Dove ~wi denota l’energia dell’i-esima autofunzione dell’Hamiltoniana H.
Infine calcolando l’integrale 1.1 si ottiene:[42]
4N X4N
1 X k2
ΦT (t) = · QT S
mn Qmn 2 (1.14)
Z m=1 n=1 k + (wm − wn )2
71 – La magnetoricezione
1.3.2 Costruzione dell’Hamiltoniana
In presenza di un campo magnetico esterno debole, a causa dell’effetto Zeeman anoma-
lo[4], si ha una perdita di degenerazione dei livelli energetici e ciò causa delle oscillazioni
tra gli stati di singoletto e di tripletto; questo comporta sia l’accentuazione dell’intercon-
versione singoletto↔tripletto sia un incremento della triplet yield.
Questo effetto ha un risultato opposto rispetto all’effetto dovuto a campi magnetici forti,
dove per forte si intende rispetto alla forza dell’accoppiamento iperfine, il quale essenzial-
mente consiste in una separazione tra i livelli energetici di stati di tripletto con numero
quantico magnetico non-nullo e l’energia dello stato di singoletto, comportando una di-
minuzione dell’interconversione singoletto↔tripletto e della triplet yield.
Osserviamo dall’equazione 1.14 che la triplet yield per campi magnetici deboli non di-
pende soltanto dalla forza del campo magnetico applicato ma anche dal rapporto tra la
forza del campo magnetico e la costante di decadimento k; per analizzare nel dettaglio
questa relazione scegliamo l’Hamiltoniana in una particolare forma semplificata, presen-
tata all’interno dell’articolo[31].
Per una coppia di radicali indicata con j, con j = 1,2, dove ogni radicale ha un singolo
elettrone con spin Sj e un singolo nucleo con spin Ij semi-intero, si ha un’Hamiltoniana
di Spin così descritta:
~ = H1 (B)
H(B) ~ + H2 (B)
~ (1.15)
con:
Hj (B)
~ = gµB S ~ + Aj I~j
~j · B (1.16)
Nell’equazione 1.16 il primo termine rappresenta l’interazione Zeeman e il secondo termi-
ne rappresenta l’interazione iperfine; µB è il magnetone di Bohr dell’elettrone.
Il fattore giromagnetico g vale 2 per entrambi i radicali.
Per campi magnetici forti (≥ 1000G), l’interconversione singoletto-tripletto può presen-
tarsi in presenza dell’interazione di Zeeman a causa della differenza nel valore di g [5],
ma questo effetto può essere trascurato in presenza di campi magnetici deboli (≤ 10G),
come quello geomagnetico, che stiamo investigando in questo caso.
Aj rappresenta il tensore di accoppiamento iperfine 3x3.
Affinché la coppia di radicali sia sensibile a diversi allineamenti rispetto al campo magne-
tico, è necessario che il tensore di accoppiamento iperfine sia anisotropico.
Nell’articolo di Ritz [31] i tensori dell’accoppiamento iperfine sono scelti nel seguente mo-
do semplificato:
Il tensore A1 per il primo radicale, scelto in modo anisotropico, è così definito:
10G 0 0
A1 = 0 10G 0 (1.17)
0 0 0
81 – La magnetoricezione
Mentre il tensore A2 per il secondo radicale, scelto invece per semplicità in modo isotro-
pico, è così definito:
5G 0 0
A2 = 0 5G 0 (1.18)
0 0 5G
1.3.3 Risultati della simulazione
In questo paragrafo presentiamo i risultati ottenuti nel calcolo dell’equazione 1.14 al va-
riare della costante di decadimento k e del campo magnetico esterno B.
Sebbene il modello scelto per l’Hamiltoniana sia particolarmente semplice, in quanto ap-
prossimiamo il comportamento biologico della coppia di radicali ad una semplice coppia
con soli due spin nucleari, assumendo che questo non cambi a causa della cinetica della
reazione, e scelti i tensori di accoppiamento iperfine come sopra citati (1.17 e 1.18), questo
non risulta essere una limitazione allo studio del modello, siccome esso serve a mostrare
come la dipendenza tra la forza dell’accoppiamento iperfine, l’intensità del campo ma-
gnetico, e il tempo di decadimento, e come l’allineamento della coppia di radicali rispetto
il campo magnetico, modifichi gli effetti del campo magnetico sulla magnetoricezione.
Dipendenza dal valore di k e di B
Il ruolo dell’intensità del campo magnetico e del tempo di decadimento sulla triplet yield
del processo di coppia di radicali con accoppiamento iperfine anisotropico è mostrato in
figura 1.3.
Figura 1.3: Andamento della triplet yield ΦT della coppia in funzione della costante di
decadimento k e dell’intensità del campo magnetico esterno B; l’angolo di allineamento
tra l’asse z della coppia e il vettore campo magnetico in questa simulazione vale 68°.
Fonte:[31]
L’asse z della coppia di radicali è stato allineato ad un angolo di 68° rispetto al vettore
campo magnetico esterno, in quanto tale angolo rappresenta, posto come vettore campo
magnetico esterno il campo geomagnetico, la più grande triplet yield. (si veda figura 1.4)
91 – La magnetoricezione
Figura 1.4: Grafico che mostra le triplet yields valutate per una coppia di radicale con
accoppiamento iperfine anisotropico. Le yields vengono mostrate sia in funzione dell’an-
golo tra il campo magnetico e l’asse z della coppia di radicali sia in funzione dell’intensità
del campo magnetico esterno. Fonte:[31]
Possiamo osservare dalla figura 1.3. che anche campi magnetici più deboli di 0,5G produ-
cono un incremento significativo della triplet yield, la quale risulta essere più pronunciata
per tempi di decadimento più grandi.
Invece per campi magnetici più intensi l’effetto è opposto: La triplet yield cala a picco.
Quest’ultimo effetto di decrescità dovuto a campi magnetici intensi fu già prevista e os-
servata sperimentalmente nei processi di coppia di radicali dai chimici Steiner e Ulrich nel
1989[40]; quest’ultimo effetto è trascurabile per lo studio della bussola aviaria, in quanto
per campi di intensità non superiori ai 5 G esso produce un cambiamento del valore della
yield trascurabile.
L’effetto delle diverse costanti di decadimento della coppia di radicale sulla triplet yields
ha un duplice aspetto:
1. Per costanti molto elevate (≥ 10µs−1 ) il veloce decadimento della coppia di radi-
cale diminuisce drasticamente l’interconversione singoletto↔tripletto, come si può
vedere dalla forte decrescita della triplet yields in assenza di campo; per costanti
di decadimento elevate l’effetto del campo magnetico esterno diventa difficilmente
osservabile.
2. Per costanti più piccole (≤ 10µs−1 ) e di campi magnetici più intensi, la forte cre-
scita della triplet yield è essenzialmente inalterata dalla costante di decadimento.
In contrasto, l’effetto di campi magnetici deboli dipende significativamente dalla
costante di decadimento.
101 – La magnetoricezione
Nella figura 1.5 sono mostrati gli effetti di campi magnetici deboli sulla triplet yield della
coppia di radicali.
Figura 1.5: la figura mostra l’influenza significativa della costante di decadimento del-
la coppia di radicali sull’intensità dell’effetto di campi magnetici deboli (come quello
geomagnetico) sulla bussola aviaria. Fonte:[31]
In conclusione è possibile affermare che la sensibilità del sistema di coppia di radicali in
presenza campi magnetici deboli cresce significativamente per costanti di decadimento
più piccole.
Quanto vale la costante di decadimento della coppia di radicali nei processi biologici?
A questo punto sorge spontaneo chiedersi quale possa essere il valore della costante di
decadimento k nei processi biologici della coppia di radicale e quindi quale possa essere
la curva che meglio descrive il fenomeno.
La costante di decadimento k del meccanismo di coppia di radicali è stata misurata in
un ambiente proteico attraverso la fotolisi a flash[28] e il risultato di tale esperimento
indica una costante di decadimento k compresa tra 1µs−1 e 2µs−1 , in base alla scelta
della proteina e del radicale in esame.
Sebbene questo risultato possa essere non considerato del tutto indicativo, mostra come
sia possibile ottenere una costante di decadimento di 1µs−1 in sistemi biologici.
Dipendenza dall’angolo θ
Per ottenere informazioni direzionali dal campo geomagnetico non basta sapere che un
campo di 0,5G influenzi la reazione di coppia di radicali, ma è necessario che ci sia una
differenza sostanziale nella triplet yield per un campo con intensità costante di 0,5G ma
con diversa orientazione rispetto all’asse z della coppia.
Riprendiamo la figura 1.4, in cui si mostra la dipendenza di ΦT da θ.
111 – La magnetoricezione
Il grafico si limita ad un intervallo di θ compreso tra 0° e 90°, in quanto la simmetria
della dinamica di spin ci permette di non considerare un intervallo angolare più ampio
siccome gli effetti orientativi dipendono soltanto dall’angolo compreso tra la direzione
del campo e un’asse z arbitrario della coppia di radicali. Inoltre, la direzione del campo
è simmetrica rispetto a un cambio di inclinazione dell’angolo di 180°; questa simmetria
deriva dalla quasi distribuzione isotropica dello spin nucleare nello stato iniziale della
coppia di radicali.[39]
Possiamo osservare dalla figura 1.4. che per un campo di 0,5 G sono possibili variazioni
di triplet yield fino al 25%; inoltre variazioni orientative non aumentano con l’aumentare
dell’intensità del campo magnetico, ciò comporta che in presenza di campi poco intensi,
a prescindere dal valore del campo geomagnetico (0,2 − 0,7 G) la variabile che "conta",
una volta fissato il valore di k e i parametri dell’accoppiamento iperfine, è l’angolo tra il
campo e l’asse z della coppia a fornire informazioni direzionali all’animale in questione.
12Capitolo 2
Visione Magnetica
A questo punto resta il problema di dove possa trovarsi nel corpo degli uccelli il mecca-
nismo della coppia di radicali.
Il posto più ragionevole al momento sembra essere nell’occhio: [21] [49]
Informazioni sulla direzione del campo geomagnetico possono essere captate dalle capa-
cità sensoriali di un’animale soltanto se il percorso di trasduzione sensoriale può essere
influenzato da una reazione che coinvolge il meccanismo di coppia di radicali e se il sistema
molecolare involto in quest’ultimo è fissato in una certa direzione spaziale. Quest’ultima
condizione può facilmente essere verificata siccome il meccanismo di trasduzione senso-
riale che concerne le membrane cellulari è spesso associata a strutture ordinate in grande
scala con determinate direzioni preferenziali definite in relazione agli organi sensoriali
dell’animale; un esempio pertinente è proprio la visione dell’animale, in quanto i coni e
i bastoncelli, ovvero i fotorecettori della retina, i quali contengono i pigmenti ottici nella
membrana, sono orientati tangenzialmente rispetto alla retina dell’occhio.
Nel seguente capitolo analizzeremo il modello di bussola aviaria basato sulla visione e i
calcoli effettuati presentati da Schulten et al nel 2000. [31]
2.1 Modello di bussola aviaria basato sulla visione
2.1.1 Prototipo dell’occhio
Per modellare l’effetto di un processo di coppia di radicali dipendente da un campo sulla
visione dell’animale è necessario specificare come il processo interagisca con il percorso
visivo.
Ai fini dell’analisi del modello assumiamo che il processo di coppia di radicali influenzi la
sensibilità dei recettori della luce nell’occhio dell’animale.
In seguito presenteremo uno schema di risposta, il quale mostra la variazione di sensibilità
su tutto l’emisfero dell’occhio.
Per semplicità rappresentiamo il nostro occhio come una fotocamera stenoscopica, ovvero
con un piccolo foro in ingresso, indicato con un il punto O’, capace di proiettare la luce,
come un obiettivo, creando un’immagine. (Si veda figura 2.1)
132 – Visione Magnetica
Figura 2.1: Prototipo dell’occhio di un pettirosso per i calcoli delle variazioni degli schemi
di risposta. Fonte:[31]
I raggi 1 e 2 entrano attraverso il foro infinitesimo O’ e sono proiettati in una retina
sferica; inoltre, assumiamo che i recettori molecolari della luce siano orientati in modo
normale rispetto alla superficie della retina (direzioni Z1 e Z2 in figura), formando angoli
diversi con la direzione del vettore campo geomagnetico.
Per facilità prendiamo in considerazione un occhio ciclopico, disposto al centro della te-
sta; la direzione della linea centrale che connette O e O’, d’ora in avanti, rappresenterà
la direzione di visione, ovvero della testa dell’animale.
2.1.2 Schemi di risposta
Definito il segnale s, rappresentato da un numero intero compreso in un intervallo tra
0 e 255, dove ad ogni numero è associata una cromaticità di nero crescente, ovvero 0 è
bianco e 255 è nero, mostriamo come varia la visione dell’animale se quest’ultimo guarda
inizialmente uno schermo uniformemente grigio, ovvero ogni pixel dell’immagine ha s =
127.
Il segnale modificato s0 è calcolato utilizzando la seguente formula di trasduzione lineare:
s0 (θ) = 127 + 255 × [(ΦT (θ) − Φgauge ) M ] (2.1)
Dove ΦT è calcolata con l’equazione 1.1 mentre Φgauge rappresenta la triplet yield mediata
su tutti gli angoli ed M è il fattore di amplificazione.
Si è scelto un valore di M = 4, in quanto questo valore del fattore di amplificazione
rende gli effetti del campo geomagnetico distinguibili per l’occhio umano in un’immagine
statica.
Nella pratica, ovvero in natura, fattori di amplificazioni molto più deboli sono sufficienti
per rendere gli effetti del campo geomagnetico distinguibili poiché un uccello può indi-
viduare la variazione dello schema visivo indotto dal campo geomagnetico in modo più
facile in quanto schemi in movimento sono riconosciuti visivamente con maggiore sensibi-
lità; in altri termini, siccome in volo l’uccello muove la propria testa si ha a che fare con
142 – Visione Magnetica
modelli in movimento e non con immagini statiche.
In ogni modo, questa maggiore sensibilità può essere illustrata soltanto mediante un vi-
deo che rappresenta la visione dell’animale, ma ciò non può essere rappresentato nella
seguente tesi, dunque limiteremo la nostra discussione ad immagini statiche.
Gli schemi visivi che descrivono la modifica della vista per diverse direzioni della testa
sono rappresentate in figura 2.2.
La numerazione da "0" a "180" si riferisce ai diversi angoli compresi tra la direzione della
testa e il vettore campo geomagnetico; si è scelto tale intervallo di valori per s e tale
scala cromatica in modo da ottenere un disco opaco nel centro di visione per lo schema di
modulazione di un uccello che guarda parallelamente alle linee del campo geomagnetico
("0" in figura 2.2).
Biologicamente, questo potrebbe corrispondere ad una diminuzione della sensibilità dei
recettori in quella regione. In ogni modo, la bussola aviaria potrebbe lavorare in modo
simile se la sensibilità dei recettori aumenta piuttosto che diminuire.
Come può essere visto nella figura 2.2, lo schema visivo per un uccello che guarda anti-
parallelamente rispetto alle linee del campo magnetico ( "180") è identico allo schema per
"0" e ciò è concorde con il fatto che la bussola aviaria basata sulla coppia di radicali sia
intrinsecamente una bussola ad inclinazione e non una bussola classica.
Siccome la bussola ad inclinazione non distingue il polo sud dal polo nord, in quanto nel-
l’emisfero boreale le linee del campo geomagnetico si inclinano verso il basso in direzione
del polo nord, mentre nell’emisfero australe le linee del campo geomagnetico si inclinano
verso il basso in direzione del polo sud, e dunque gli schemi visivi sono indistinguibili,
tale informazione viene fornita all’uccello grazie al suo sistema vestibolare situato nell’o-
recchio interno.[30]
In ogni modo sorge lo stesso problema nel momento in cui l’animale vola in prossimità
dell’equatore e perde la capacità vestibolare di differenziare il polo nord dal polo sud;
limitandoci all’analisi del pettirosso, questo problema è poco vincolante in quanto tale
animale migra dall’Europa al Nord Africa, restando "lontano" dall’equatore.
Per capire come interpretare le informazioni ricevute dal campo geomagnetico ottenute
dal meccanismo di coppia di radicali, bisogna considerare come questa informazione ap-
paia durante il movimento della testa:
Se l’uccello vola parallelamente rispetto al vettore campo magnetico e muove la testa
all’insù, il disco opaco seguirà il movimento dei suoi occhi con una velocità angolare
ridotta; In effetti la velocità angolare dipende dalla geometria della lente oculare. Nel
nostro modello, il disco si muove con una velocità angolare che è la metà della velocità
angolare dell’occhio, in quanto come è osservabile dalla figura 2.1 la variazione angolare
del raggio 1 è la metà di quella di Z1 , siccome l’angolo (1, 0’, C) è la metà dell’angolo
(1, O, C). Se l’uccello facesse su e giù con la testa, vedrebbe il disco salire e scendere
seguendo il movimento della testa e tale disco risulterebbe centrato soltanto se l’animale
guarda parallelamente o anti-parallelamente rispetto alla direzione del campo magnetico.
Ragionamento analogo per il movimento laterale destra sinistra della testa.
I pittogrammi in figura 2.2 non rappresentano comunque situazioni comportamentali da
parte dell’uccello realistiche, in quanto in volo tali animali cercano di stare con la testa
in posizione orizzontale per facilitare il volo.
152 – Visione Magnetica
Figura 2.2: Schemi di modulazione visivi dovuti al campo geomagnetico 0,5 G per un
uccello che guarda in direzioni distinte ad angoli di 0°, 30°, 60°, 90°, 120°, 150°, e 180°
rispetto al vettore campo geomagnetico. Le illustrazioni schematiche vicino agli schemi
visivi indicano la corrispondente direzione lungo la quale viaggia l’animale ad Urbana-
Champaign (linea di campo geomagnetico inclinata di 68°). Fonte:[31]
Per rappresentare un esempio più realistico, mostriamo nelle figura 2.3 e 2.4 come gli
schemi visivi cambiano se l’uccello ruota in un piano parallelo alla superficie terrestre;
la rotazione in questo piano comporta una sovrapposizione degli effetti discussi in prece-
denza, ovvero una sovrapposizione dello spostamento della testa laterale e verticale.
Nella figura 2.5 vengono mostrati gli schemi di modulazione visivi per campi di diversa
intensità con l’animale che guarda parallelamente rispetto alle linee del campo.
La diversa intensità del campo ha effetti sul contrasto e sulla forma degli schemi visivi.
Il contrasto è più forte per campi di 0,5 G e 1,0 G e diminuisce per campi più o meno
intensi, risultando appena percettibile per campi di 0,1 G.
Questi risultati sono compatibili con le evidenze sperimentali ottenute da Wiltschko et al
nel 1972 [50] come sopra citate.
Inoltre questo modello riesce a spiegare ulteriori osservazioni sperimentali inizialmente
non comprensibili [49]; Wiltschko osservò che dopo 3 giorni di acclimatamento gli uccelli
erano capaci di orientarsi anche in presenza di campi con intensità maggiori o minori del
intervallo precedentemente citato, ad esempio 0,16 o 1,5 G; inoltre in caso di acclimata-
mento per un campo di 1,5 G, gli uccelli erano in grado di orientarsi sia per il campo di
1,5 G sia per quello geomagnetico di 0,5 G, ma non per campi di intensità intermedi, ad
esempio di 0,8 G.
Gli schemi 2.4 ci permettono di fornire una possibile spiegazione a queste osservazioni:
per campi molto intensi cambia soltanto la forma del disco e il suo contrasto, dopo diver-
si giorni l’animale riesce ad abituarsi al nuovo schema e dunque a ricavare nuovamente
informazioni dal nuovo campo magnetico, senza perdere la capacità di orientarsi per il
campo precedente.
162 – Visione Magnetica
Figura 2.3: Schemi di modulazione visivi dovuti al campo geomagnetico per un uccello
che vola parallelamente all’orizzonte ad Urbana-Champaign e guarda in diverse direzioni.
Fonte:[31]
Figura 2.4: Visione panoramica di Francoforte, Germania. L’immagine mostra la prospet-
tiva del paesaggio registrata da un uccello che vola ad altitudine di 200 metri rispetto al
suolo e guarda in direzione dei vari punti cardinali. L’inclinazione del campo magnetico
è di 66°, valore medio di quella regione. Fonte:[38]
172 – Visione Magnetica
Figura 2.5: Schemi di modulazione visivi per campi magnetici di diverse intensità per un
uccello che guarda in direzione parallela al vettore campo magnetico. Fonte:[31]
Osservazioni
Gli schemi di modulazione visivi presentati in questo capitolo devono essere considerati
formali, delle rappresentazioni antropomorfiche degli stimoli dell’occhio dell’animale at-
traverso un sistema ordinato di fotorecettori dovuti a campi magnetici statici esterni, e
non quello che effettivamente il volatile vede. Siccome il reale apparato visivo del petti-
rosso contiene due occhi, fornendo un immagine a 360°, e presenta variazioni di sensibilità
su diverse aree della retina e una geometria della retina non perfettamente sferica, com-
porterà ulteriori variazioni degli schemi di modulazione visivi e differenze nel modo in cui
questi elaborano le informazioni. Ulteriori modelli in cui si tiene conto di queste correzioni
non verranno analizzati nella seguente tesi.
2.2 I candidati magnetorecettori
A questo punto bisogna trovare un candidato magnetorecettore per la bussola aviaria.
2.2.1 La magnetite
In questo paragrafo presenteremo inizialmente, per completezza di discorso, l’ipotesi sulla
quale fino a pochi anni fa si pensava si potesse basare la magnetoricezione di alcuni
animali, ovvero la presenza di cristalli di magnetite, un minerale con naturali proprietà
magnetiche, in molti degli animali e microrganismi che sembravano possedere questo senso
magnetico.
L’ipotesi che la magnetoricezione potesse basarsi sulla presenza di cristalli di magnetite
all’interno del corpo degli animali nacque dall’idea che il senso magnetico biologico della
bussola aviaria potesse assomigliare ad una classica bussola usata dall’uomo, ovvero un
ago magnetico formato dalla magnetite (F e3 O4 ), il quale presenta risposte classiche in
presenza di un campo magnetico esterno.
182 – Visione Magnetica
In particolare nei primi anni del duemila alcune ricerche mostravano che i cristalli di
magnetite sembravano trovarsi in certi neuroni all’interno del becco superiore dei piccioni
viaggiatori [13], e questo suggeriva che i loro neuroni rispondessero a segnali magnetici
raccolti dai cristalli e poi mandati a loro volta nel cervello dell’animale; inoltre ulteriori
evidenze sperimentali mostravano che i piccioni si disorientano e perdono la loro abilità
di rilevare il campo geomagnetico quando hanno attaccate al becco delle piccole calamite,
dove pare si trovino i neuroni contenenti magnetite.[29]
In effetti, dal punto di vista classico, un piccolo cristallo di magnetite con un diametro
di circa 50 nm ha un momento magnetico permanente grande abbastanza da permettere
l’allineamento del cristallo con un campo magnetico di bassa intensità, anche di 0,5 G,
fornendo informazioni direzionali alla bussola aviaria. [19][18]
Se ci fosse una catena di piccole particelle di magnetite, il momento magnetico indotto
causerebbe un’attrazione e repulsione tra i cristalli adiacenti causando un allungamento
o accorciamento della catena, effetto che verrebbe rilevato da un eventuale recettore
meccanico e che rappresenterebbe la base della risposta del senso magnetico alla variazione
dell’intensità del campo magnetico esterno.
Tuttavia, sebbene sembrava si fosse finalmente individuato il sito della magnetoricezione,
si dovette rincominciare tutto da capo nel 2012 quando un gruppo di ricerca australiano
pubblicò un articolo[44] in cui si descriveva un particolareggiato studio in tre dimensioni
del becco dei piccioni, usando la risonanza magnetica, e si concludeva che quelle cellule,
chiamate macrofagi, contenenti magnetite nel becco superiore dell’uccello certamente non
hanno nulla a che fare con la magnetoricezione, ma difendono l’animale da infezioni,
ingerendo microrganismi invasori o rimuovendo cellule morte per apoptosi.
2.2.2 I crittocromi
Note le ipotesi e i risultati presentati nel modello precedente, possiamo elencare i criteri
essenziali che deve avere un candidato magnetorecettore:
1. Il magnetorecettore deve essere legato biologicamente ad un fotorecettore (solo luce
blu o verde) che inizia il processo di coppia di radicale.
2. Il magnetorecettore deve contenere una coppia di molecole capaci di compiere rea-
zioni chimiche basate sul meccanismo di coppia di radicali influenzato da un campo
magnetico debole.
3. Siccome abbiamo supposto che tale processo avvenga nell’occhio, il magnetorecetto-
re deve essere connesso alla catena di trasduzione nervosa visiva; inoltre i recettori
devono essere disposti in un modo ordinato in modo tale da fornire la dipendenza
dall’orientazione necessaria affinchè funzioni la bussola aviaria; ad esempio la loro
orientazione deve ricoprire un vasto intervallo angolare.
Attualmente si suppone che i magnetorecettori ricercati siano i crittocromi; i crittocromi
sono una classe di flavoproteine, cioè contenenti una molecola derivata dall’acido nuclei-
co riboflavina( il flavina adenina dinucleotide FAD), responsabili della sincronizzazione,
scatenata dalla luce, dei ritmi circadiani di molte specie viventi [26].
192 – Visione Magnetica
In seguito elenchiamo in che modo tali composti organici soddisfano i requisiti sopra
elencati:
1. I crittocromi possiedono al proprio interno fotorecettori di luce blu, chiamate flavine
(FAD), che avviano il processo di coppia di radicale. [1] [7].
2. Dopo aver assorbito un fotone di luce blu, il pigmento flavina adenina dinucleotide
FAD emette un elettrone da uno dei suoi atomi, lasciando al suo posto una lacuna
elettronica; questa può essere occupata da un altro elettrone , donato da una coppia
di elettroni in correlazione appartenenti ad un amminoacido, chiamato triptofano,
conenuto sempre all’interno della proteina crittocromo. Questo processo corrisponde
al meccanismo di coppia di radicali ricercato. [22]
3. I crittocromi sono stati scoperti sulla membrana limitante interna della retina
di diversi animali (ad esempio i topi [26]) sulla quale risultano essere distribuiti
uniformemente presentando appunto un grande intervallo angolare di orientazione.
Nel 2004 tre tipi di crittocromi sono stati trovati negli occhi dei pettirossi.[27]
20Capitolo 3
Verifiche sperimentali
In questo capitolo presenteremo l’esperimento eseguito da Ritz et al a Francoforte [32], il
quale ci permette di identificare il processo di coppia di radicali come il candidato prin-
cipale per il funzionamento della bussola aviaria, mostrando ulteriormente come l’ipotesi
della magnetite è inadatta a spiegare tale fenomeno.
A conclusione di questo progetto di tesi analizzeremo brevemente alcuni problemi e aspetti
ancora incompresi della magnetoricezione.
3.1 L’esperimento di Ritz
L’esperimento eseguito da Ritz et al mostra come campi magnetici oscillanti disturbano
la capacità degli uccelli di orientarsi, e aumentare o diminuire la frequenza del campo lo
rende meno efficiente nella sua azione di disturbo.
Questo esperimento stabilisce inoltre come campi magnetici oscillanti possono essere usati
come strumenti per stabilire l’effettivo coinvolgimento di una reazione di coppia radicale
negli uccelli; in aggiunta studiando l’intervallo di frequenze all’interno del quale campi
oscillanti modificano il meccanismo di coppia di radicale, si può pervenire ad ulteriori
informazioni riguardo la natura chimica della coppia di radicali coinvolta.
3.1.1 Modello matematico
Per calcolare teoricamente la veridicità dell’esperimento si è ricalcolata la triplet yield
(si veda equazione 1.2) in assenza e presenza di campi oscillanti, mostrando come per
ottenere la stessa variazione della triplet yield dovuta a campi oscillanti poco intensi, è
necessario un campo magnetico molto più intenso di quello oscillante.
Per fare ciò, si è scelto inizialmente un modello di coppia di radicali con un solo protone
e con accoppiamento iperfine, ignorando l’accoppiamento dipolare tra gli elettroni ed
omettendo l’iterazione di Zeeman in quanto ci interessano campi magnetici molto deboli.
In termini dell’Hamiltoniana del sistema questa corrisponde all’equazione 1.16 con un
solo protone, ovvero: [43]
Ĥ = aŜR · Iˆ + ŜR · A · Iˆ + ω ŜRz + ŜPz
(3.1)
213 – Verifiche sperimentali
Dove con R si indica il radicale contenente il protone; a è la parte isotropica dell’interazio-
ne iperfine; A è un tensore a traccia nulla contenente la parte anisotropica dell’interazione
iperfine; ŜR è l’operatore momento angolare di spin elettronico per il radicale R e ŜRz è
la sua componente lungo la direzione del campo magnetico (posto lungo l’asse z); ŜP è
l’operatore momento angolare di spin protonico per il radicale R; Iˆ è l’operatore momento
angolare di spin nucleare; La frequenza di Larmor ω dello spin elettronico è calcolato nel
seguente modo ω = gµB B0 /~, dove si è scelto il fattore giromagnetico uguale a 2 per
entrambi gli elettroni.
Indicate con αx , αy e αz le componenti non nulle diagonali di A, definiamo l’assialità del
tensore nel seguente modo:
(aX + aY )
α= (3.2)
2a
Nel calcolo teorico presentato nel seguente esperimento si sono scelti i seguenti parametri:
a di 5 G, α di 0,3, ed un tempo di vita di 20µs (tempo di vita medio della flavina-triptofano
osservato sperimentalmente nella coppia di radicali [14]).
Dopo aver risolto l’equazione di Liouville-von Neumann (equazione 1.7) in presenza di
un campo magnetico statico di 0,46 G, si è determinato, mediante integrazione numerica
dell’equazione 1.7, la variazione nella triplet yield ΦO causata da un campo magnetico
oscillante di 1,3 MHz in risonanza con lo splitting dovuto al campo statico di 0,46 G;
per confronto, calcolando la variazione della triplet yield dovuta ad una diminuzione
del campo magnetico statico ΦS a 0,12 G, in quanto si è notato sperimentalmente che
ciò comporta un disorientamento dell’animale [50], tale variazione risulta essere uguale
a quella ottenuta per un campo oscillante di 1,3 MHz con intensità di 0,3 mG, ovvero
molto meno intenso dei campi presenti nell’esperimento.
3.1.2 Apparato strumentale e procedura del test
L’esperimento è stato eseguito in capanne di legno nel giardino dell’istituto Zoologico
di Francoforte, dove il campo geomagnetico locale presenta un intensità di 0,46 G e
un inclinazione di 66° rispetto la direzione Nord, utilizzando 12 pettirossi catturati in
autunno durante la loro migrazione verso sud ed il test è stato eseguito in primavera
quando questo animale emigra verso nord.
Per generare campi oscillanti è stata utilizzata una bobina (210 cm di diametro) montata
su un telaio di legno girevole che circondava la zona del test.
Gli uccelli sono stati posti in degli imbuti di Emlen e venivano testati individualmente
una volta al giorno.
Un imbuto di Emlen è fatto nel seguente modo: è un imbuto a forma di cono rovesciato
(nel seguente esperimento il diametro massimo superiore misura 35 cm, mentre l’altezza
20 cm) con del tampone inchiostrato sul fondo e carta assorbente sull’interno dei lati
inclinati (si veda figura 3.1); quando un uccello salta, o svolazza sui lati dell’imbuto,
lascia tracce che danno informazioni sulla direzione che cerca di prendere il volatile per
volare via. La sommità dell’imbuto presenta un emettitore di luce blu/verde (il picco di
frequenza è a lunghezza d’onda λ = 565nm e il picco a mezza altezza è a una lunghezza
d’onda di 533 e 583 nm) che raggiunge l’uccello con un intensità di 2.1 mWm−2 .
223 – Verifiche sperimentali
Il test iniziava quando la luce viene emessa nelle gabbiette e durava 75 minuti ogni volta.
Ogni uccello è stato testato 3 volte in ogni condizione ma con un ordine casuale, in modo
tale da non poter creare eventuali condizioni di acclimatamento per l’animale.
Figura 3.1: Esempio di tubo di Emlen. Fonte:[2]
3.1.3 Analisi dei dati
Per l’analisi dei dati si è divisa la carta assorbente nell’imbuto di Emlen in 24 settori,
e le tracce rilasciate dalle zampette degli uccelli in ogni settore sono state contate da
sperimentatori ignari dell’indagine in corso, in modo tale da non influenzare l’esito della
raccolta dati.
Ricavate le tre direzioni di volo dell’uccello per ogni condizione del test, si è calcolato il
vettore medio con lunghezza rb e angolo αb per ogni condizione.
I dati raccolti sono mostrati nella tabella 3.3.
Con i 12 vettori costruiti si è ottenuto per ogni condizione un Grand mean vector uti-
lizzando il test di Rayleigh (si veda appendice A), in modo tale da ricavare informazioni
sulla dispersione direzionale degli uccelli in ogni condizione.
I test sono stati eseguiti nelle seguenti diverse condizioni:
a. Presenza del solo campo geomagnetico.
b. Presenza del campo geomagnetico e di un campo oscillante di disturbo a banda larga
(0.1–10 MHz) con intensità di 0,85 mG, in quanto molte biomolecole presentano lo
splitting dovuto all’interazione iperfine in quel range di frequenze.[43]
c. Presenza del campo geomagnetico e di un campo di disturbo di 4,7 mG con frequen-
za di 7,0 MHz posto parallelamente rispetto il campo geomagnetico (24° rispetto al
nord geografico in senso orario).
d. Presenza del campo geomagnetico e di un campo di disturbo di 4,7 mG con frequen-
za di 7,0 MHz posto ad un angolo di 24° rispetto il campo geomagnetico (Parallela
rispetto al nord geografico).
e. Presenza del campo geomagnetico e di un campo di disturbo di 4,7 mG con fre-
quenza di 7,0 MHz posto ad un angolo di 48° rispetto il campo geomagnetico (24°
rispetto al nord geografico in senso antiorario).
233 – Verifiche sperimentali
Una rappresentazione grafica delle direzioni medie di volo degli uccelli è mostrata nella
figura 3.2.
Figura 3.2: Effetti di campi magnetici oscillanti sul comportamento direzionale dei petti-
rossi. I triangolini indicano la direzione di volo media dei 12 uccelli nelle varie condizioni.
Le freccie rappresentate indicano i grand mean vectors. Fonte:[32]
Figura 3.3: Tabella dei dati raccolti durante l’esperimento di Ritz; con rb e αb si intende
rispettivamente la lunghezza e l’angolo del vettore direzionale del volo dell’uccello rispetto
alla direzione nord della circonferenza ottenuto per ogni condizione del test.
Nell’ultima riga sono riportati i Grand mean vectors e l’angolo medio ottenuti per ogni
condizioni. Fonte:[32]
243 – Verifiche sperimentali
3.1.4 Risultati
Come si evince dai dati raccolti mostrati nelle figure 3.2 in condizioni normali (figura
3.2a) i pettirossi mostrano un’orientazione di volo diretta verso nord, come ci si aspetta
dalle loro classiche migrazioni primaverili, ma in presenza di un campo di disturbo oscil-
lante l’animale risulta fortemente disorientato (figura 3.2b).
Per confermare che la modifica comportamentale dell’animale è causata da un effetto
diretto del campo magnetico oscillante sulla bussola aviaria e non da effetti aspecifici, si è
variato l’allineamento del campo ad una frequenza di 7.0 MHz: come si osserva dalle figure
3.2c,3.2d e 3.2e, quando il campo oscillante è posto parallelamente al campo magnetico
gli uccelli risultano essere ben orientati nelle classiche direzioni di migrazione e non c’è
una differenza osservabile dal loro classico comportamento; in contrasto, quando lo stesso
campo oscillante viene posto ad un angolo diverso, rispettivamente 24° e 48° rispetto al
campo geomagnetico, gli uccelli risultano essere fortemente disorientati.
Le frequenze per le quali si mostra una perturbazione dovuta a campi oscillanti per il
meccanismo di coppia di radicali non dipendono soltanto dalla natura chimica della cop-
pia, ma anche dall’allineamento del campo oscillante rispetto al campo statico (quello
geomagnetico in tal caso.).[6]
Questo implica che siccome la risposta della bussola aviaria basata sul meccanismo di
coppia di radicali può dipendere dall’allineamento tra il campo oscillante e il campo geo-
magnetico, ulteriori effetti aspecifici di disturbo sul comportamento dell’animale risultano
essere indipendenti da quest’ultimo.
Grazie a questo esperimento, inoltre, è possibile escludere l’ipotesi che la magnetoricezio-
ne nei pettirossi possa avvenire grazie alla magnetite, in quanto non sono previsti cambi
di allineamento magnetico per recettori di magnetite per campi oscillanti così deboli fino
a frequenze più grandi di 100 kHz[17].
3.2 Aspetti irrisolti
Questi studi hanno stimolato un’esplosione di interesse nella magnetoricezione, spingendo
tanti ricercatori a trovare altri animali che possano possedere questo senso magnetico,
addirittura un’equipe della University of Auckland studiando le migrazioni della trota ha
osservato che quest’animale possiede la magnetoricezione; inoltre studiando l’anatomia
dell’animale si è ipotizzato che il luogo in cui potrebbero trovarsi i magnetorecettori possa
essere il naso.
Se l’ipotesi fosse corretta, sarebbe il primo esempio di una specie che riesce ad annusare
il campo magnetico terrestre.[46]
Nonostante si siano fatti incredibili passi avanti nella comprensione del fenomeno, la
scoperta della magnetoricezione in diverse specie di animali ha portato alla nascita di
diverse questioni alle quali ancora oggi non si giunge ad una conclusione univoca; per
esempio, come fanno i radicali liberi della coppia a rimanere in correlazione quantistica
sopravvivendo alla decoerenza abbastanza a lungo da fare la differenza dal punto di vista
biologico.
25Puoi anche leggere

















































